1. Вторичные сукцессии. Демутации.
2. Ассоциация как низшая и основная единица классификации.
3. Местообитание, экотоп, биотоп.
4. Горизонтальная структура биоценоза. Мозаичность. Микроценозы.
5. Вторичные сукцессии. Пастбищные дигрессии.
6. Численность видов в биоценозе. Доминанты, субдоминанты, второстепенные виды.
7. Учение В. И. Вернадского о биосфере.
8. Видовой состав биоценоза. Видовая насыщенность. Изменения видового разнообразия по градиентам среды.
9. Экологические области океана.
10. Живое вещество биосферы и его химический состав.
11. Реликт, реликтовый ареал.
12. Понятие “климакс”. Представление об устойчивости биоценозов.
13. Понятия “биоценоз”, “фитоценоз”, “животное население”. Основные подходы к их изучению.
14. Ценотическая значимость видов. Эдификаторы, соэдификаторы, ассектаторы.
15. Наиболее общие закономерности сукцессий. Смена жизненных форм, видового разнообразия, изменение биомассы.
16. Зональное расчленение живого покрова суши. Важнейшие градиенты среды.
17. Сообщества организмов океана.
18. Флуктуации биоценозов.
19. Первичные сукцессии. Стадии формирования биоценоза.
20. Роль конкурентных отношений между видами и организмами биоценоза. Внутривидовые взаимоотношения.
21. Круговорот кислорода.
22. Пресные водоемы как среда жизни.
23. Подходы к выделению жизненных форм. Жизненные формы животных.
24. Межвидовые взаимоотношения в биоценозе. Принцип конкурентного исключения и дифференциации экологических ниш.
25. Представление о континууме. Границы биоценозов.
26. Биоценоз и условия местообитания. Средообразующая роль живых организмов.
27. Биоценоз, биогеоценоз, экосистема.
28. Жизненные формы растений. Классификация К. Раункиера.
29. Факторы, обуславливающие верхний предел жизни в горах. Специфика высокогорных биоценозов.
30. Жизненные стратегии видов. Фитоценотипы Л. Т. Раменского.
31. Соленость как экологический фактор в распространении морских организмов.
32. Центры таксономического разнообразия в ареале.
33. Биогеографические области Мирового океана.
34. Понятия “флора”, “фауна”. Основные подходы к их изучению.
35. Ареал. Ареалы космополитные и эндемичные.
36. Вертикальная структура биоты Мирового океана.
37. Сезонная изменчивость биоценозов. Смена аспектов.
38. Биогенные круговорот углерода (С).
39. Границы ареалов и факторы, их определяющие.
40. Вертикальная структура биоценоза. Ярусность наземная и подземная.
41. Первичные сукцессии. Стадии формирования биоценозов.
42. Биогенный круговорот азота (N).
43. Флуктуации биоценозов (см. 18).
44. Границы ареалов видов и факторы, их определяющие (см. 39).
45. Понятие “структура ареала”.
46. Моря и океаны как среда жизни.
47. Развитие ареалов во времени. Дизъюнктивные ареалы.
48. Высотная поясность в горах. Понятия “высотный пояс”, “тип поясности”.
49. Вторичные сукцессии. Пирогенные сукцессии.
50. Эндемики. Палео- и неоэндемики.
51. Демутации и дигрессии.
52. Антропогенные сукцессии, их важнейшие особенности.
53. Охрана редких видов животных и растений. “Красные книги”.
54. Эндемики и эндемичные ареалы.
55. Эндоэкогенетические (автогенные) сукцессии.
56. Экзоэкогенетические (аллогенные) сукцессии.
57. Зональные, интра- и экстразональные биоценозы. Их место в структуре растительного покрова зоны.
58. Климакс. Представление об устойчивости биоценозов (см. 12).
59. Роль человека в формировании ареалов.
60. Основные различия между биоценозами песчаных и глиняных пустынь.
61. Пелагиаль как экологическая область океана.
62. Голарктическое флористическое царство. Восточно-азиатская область.
63. Важнейшие изменения растительности и животного населения тайги с севера на юг.
64. Характеристика листопадных тропических лесов.
65. Важнейшие особенности тундровых биоценозов. Жизненные формы, структура, изменчивость.
66. Изменение состава и структуры степных биоценозов с севера на юг.
67. Географические особенности размещения темнохвойных и светлохвойных лесов России.
68. Неотропическое флористическое царство. Амазонская и Андийская области.
69. Характеристика биома саванн.
70. Растительность и животное население степей.
71. Важнейшие особенности биоценозов темнохвойных таежных лесов.
72. Важнейшие особенности организации пустынных биоценозов.
73. Неотропическое флористическое царство. Амазонская и Андийская области (см. 68).
74. Характеристика степного биома.
75. Характеристика биоценозов тропических влажных лесов.
76. Краткая характеристика Австралийской фаунистической области.
77. Растительность и животное население листопадных тропических лесов.
78. Капское флористическое царство.
79. Растительность и животное население саванн.
80. Растительность и животное население биома широколиственных лесов.
81. Характеристика Эфиопской области.
82. Сравнительная характеристика биоценозов тайги и хвойно-широколиственных лесов.
83. Сравнительная характеристика биоценозов тундры и лесотундры.
84. Сравнительная характеристика биоценозов степи и лесостепи.
Ответы на вопросы
2. Ассоциация как низшая и основная единица классификации
Один из наиболее широко применяемых принципов построения таксономических категорий, основанный на выделении доминантов и эдификаторов, лучше других приложим к выделению таксономических категорий биоценозов. Этот принцип является в то же время физиономичным, т. к. сообщества с экологически близкими доминантами имеют и сходный облик.
Построенная по доминантам и эдификаторам система таксономических категорий сообществ выглядит следующим образом. Наименьшей таксономической категорией является ассоциация, объединяющая участки земной поверхности с сообществами, в наибольшей степени сходными по строению, т. е. с одинаковыми эдификаторами и доминантами, одинаковой структурой, одинаковым набором консорций. Иными словами, биоценотическая ассоциация представляет собой и совокупность сходных сообществ, и тип биоценоза. Пример ассоциации: лес с сосновым древостоем и наземным лишайниковым покровом – “бор беломошник”.
В. В. Алехин отмечал, что при познании ассоциаций основным объектом являются конкретные участки растительного покрова, связанные с особенностями рельефа, почвы, иногда биотических факторов.
Типологическими классификациями более высокого ранга являются группа ассоциаций, формация, группа формаций, класс формаций, тип биома, или биоценотический тип. К одной группе биоценотических ассоциаций относят ассоциации, которые различаются только по одному из второстепенных ярусов. Формации отличаются друг от друга по составу эдификаторов и доминантов главного яруса и т. д.
3. Местообитание, экотоп, биотоп
Формирование растительных сообществ протекает при постоянном взаимном влиянии растений, их составляющих и среды обитания. Каждое сообщество связано с определенным местообитанием. При изменении условий жизни, факторов внешней среды в процессе исторического развития сообщества, изменялись и виды, слагающие сообщества, подбирались, устанавливались и закреплялись их сочетания.
Совокупность условий, необходимых для существования того или иного вида, называется стацией (для растений – местообитание). Участок земной поверхности, характеризующийся определенными абиотическими условиями, соответствующий одному биоценозу, называется биотопом. Участок земной поверхности, характеризующийся определенными абиотическими условиями, соответствующий экосистеме любого ранга, называется экотопом.
Любой биоценоз изменяет среду обитания, с которой он связан. Сообщество характеризуется своей внутренней средой, отличающейся от среды, не преобразованной сообществом.
4. Горизонтальная структура биоценоза. Мозаичность. Микроценозы
Горизонтальная структура биоценоза мозаична. Однородность биотопа может нарушаться наличием наноформ рельефа, приводящих к различным условиям увлажнения и засоления, а следовательно, к различиям в растительном покрове. В самом биоценозе может быть некоторая неоднородность среды из-за чередования крон и подкроновых пространств, неоднородности освещенности и влажности. Различные породы, входящие в состав древесного полога могут создавать некоторую пестроту в условиях существования более низких ярусов, вызывающую различия в их составе.
Мозаичность биоценоза выражается в его подразделении на несколько типов участков – микроценозов, каждый из которых характеризуется определенным количественным соотношением видов, структурными особенностями. Каждый тип микроценоза включает организмы различного систематического положения и охватывает все ярусы биоценоза.
Основным критерием различия микроценозов одного биоценоза и двух биоценозов являются наличие или отсутствие одного (или нескольких) общих эдификаторов.
5. Вторичные сукцессии. Пастбищные дигрессии
Вторичная сукцессия – смена одного сообщества другим в результате изменений условий существования или значительных нарушениях состава сообщества.
Вторичные сукцессии могут происходить путем постепенного изменения сообщества в результате изменения им среды обитания или под влиянием внешних воздействий, не связанных с эти логическим развитием. Последний тип смен называется стихийным или внезапным. Внезапные смены могут быть: климатогенными, эдафтогенными, биогенными, пирогенными и антропогенными. Любая смена, начавшаяся под влиянием внешнего по отношению к сообществу фактора, постепенно переходит в эндодинамическую смену, определяемую все в большей степени воздействием сообщества на среду его обитания.
Внезапные смены происходят под влиянием катастрофических воздействий самого различного характера. Каждая такая смена начинается гибелью видов, характерных для данного сообщества, затем происходит постепенно восстановление этого сообщества. Любая сукцессия, вызванная катастрофической причиной, состоит из нескольких быстро сменяющих друг друга начальных стадий после произошедшей катастрофы, а затем значительно более медленное длительное восстановление исходного сообщества.
Распашка степных пространств и выращивание не месте распаханных степей зерновых культур приводит к резкому обеднению фауны. Значительная часть видов-зеленоядов исчезает. Убежища многих степных животных при распашке разрушаются, и они становятся легкой добычей врагов. Лишь немногие виды (хомячки), для которых в посевах находится пища, увеличивают свою численность, остальные сокращаются и исчезают. Если пашни спустя некоторое время забрасываются под залеж (для восстановления плодородия), начинается медленное восстановление степных сообществ. Сначала господствуют бурьянистые растений и животные – потребители семенных кормов (при значительной бедности видового состава). Вторая стадия – сообщества с преобладанием корневищных злаков, развивающиеся на некотором уплотнении почвы. В составе сообщества снова появляются норники-зеленояды. Восстанавливается состав степняков – насекомых и птиц. Происходит постепенно вселение степных видов растений – представителей разнотравья, а затем дерновинных злаков, характерных для степных сообществ птиц открытых пространств (жаворонок, степной орел), а также степных насекомых (полужесткокрылые, прямокрылые). Процесс восстановления степного сообщества длится многие десятилетия.
6. Численность видов в биоценозе. Доминанты, субдоминанты, второстепенные виды
Одна из основных особенностей любого биоценоза – его видовой состав. Общее число видов растений и животных, свойственных биоценозу данного типа, более или менее постоянно. Наиболее богаты по видовому составу биоценозы влажных тропиков, наиболее бедны – биоценозы аридных и холодных регионов. Чем больше видов в биоценозе, тем более он устойчив, т. к. в нем формируется больше связей.
Количество видов, приходящихся на единицу площади, называется видовой насыщенностью. Она изменяется в различных биоценозах в том же направлении, что и видовой состав. Видовая насыщенность различных систематических групп растений и животных в одном и том же биоценозе резко различается. Для каждого биоценоза характерен определенных набор и преобладание тех или иных групп жизненных форм.
Не все виды играют в биоценозе одинаковую роль. Обычно среди растений различают компоненты-многолетники, постоянно присутствующие в биоценозе, и ингредиенты-однолетники, количество которых может резко колебаться год от года в зависимости от условий погоды.
Среди организмов, образующих биоценоз, различают доминанты, т.е. виды преобладающие по численности. Разным систематическим группам организмов, образующих биоценоз, свойственны свои доминанты. Кроме доминантов выделяют также субдоминанты, второстепенные и третьестепенные (редкие) виды. Критерии разделения на эти группы как правило численные: число особей, биомасса, проективный покров (у растений), обилие. Некоторые ученые выделяют доминанты-растения только для главного яруса, считая господствующие растений второстепенных ярусов субдоминантами. Часто выделяют доминанты групп ярусов и даже отдельных ярусов.
По количеству доминант биоценозы бывают моно-, олиго- и полидоминантные. Часто происходит смена доминант от года к году, вызванная колебаниями параметров внешней среды (“защитная реакция биоценоза”).
7. Учение В. И. Вернадского о биосфере
Биосфера – земная оболочка, занятая совокупностью организмов, населяющих Землю. Понятие о биосфере было введено австрийским ученым Э. Зюссом в 1875 г. Стройное учение о биосфере было развито В. И. Вернадским, положившим начало ее геохимическому направлению.
Биосфера включает: 1) тропосферу, где активная жизнь может существовать до высоты 10-15 км, перенос покоящихся зачатков происходит до высоты свыше 20 км, т.е. уже в стратосфере; 2) гидросферу, в которой жизнь проникает до наибольших глубин Мирового океана; 3) верхнюю часть литосферы – кору выветривания (мощность 30-60, до 200 м). Кора выветривания – совокупность геологических отложений, образованных продуктами разложения и выщелачивания горных пород различного состава, оставшуюся на месте возникновения или перемещенную на небольшое расстояние, но не утратившую связь с материнской породой. За пределами коры выветривания жизнь может быть обнаружена только в отдельных случаях.
В. И. Вернадский указывал, что под биосферой лежит область осадочных пород (до 5-6 км) биогенного происхождения, не образующая сплошного покрова – стратисфера. Эти породы были созданы в гидросфере под действием трех главных агентов: организмов, воды и ветра (перерабатывает и перемещает породы после их поднятия над уровнем воды). В пределах биосферы существуют области, в которых активная жизнь не возможна (верхние области тропосферы, очень холодные или очень жаркие районы з.ш., где организмы могут существовать только в покоящемся состоянии). Совокупность таких областей биосферы называется парабиосферой (Хатчисон).
Вернадский выделял непрерывный слой живого вещества, занимающий водную толщу и узкой полосой простирающийся по границе литосферы и тропосферы, где он включает почву и подпочву с находящимися в них корнями растений, грибами, микроорганизмами и почвенными животными, и приземную часть тропосферы, в которой располагаются надземные части растений и переносится основная масса их пыльцы, спор и семян. Этот слой называют фитосферой или фитогеосферой, т.к. в нем основными накопителями энергии являются растения. Мощность фитосферы велика только в области океанов, на суши ее мощность не больше 100-150 м. При этом в тропосфере в отрыве от жидкой и твердой оболочек живые вещества могут находятся линь временно, т.к. некоторые функции, например размножение, здесь не могут быть осуществлены.
Хатчисон выделяют следующие три особенности биосферы:
1. В значительных количествах содержится жидкая вода.
2. Падает мощный поток энергии солнечных лучей.
3. Находятся поверхности раздела между веществами, находящимися в трех фазах: твердой, жидкой и газообразной.
Биомасса животных равна приблизительно 1016 г, биомасса растений – 1019 г. На суше биомасса растений примерно на три порядка больше биомассы животных, в океане биомасса животных в 28 раз больше (но очень велика продуктивность растений на единицу площади – в 200 раз больше, чем на суше). Общая биомасса океана на три порядка меньше биомассы суши.
Масса биосферы незначительна по сравнению с массой Земли, однако при такой массе организмы осуществляют свою планетарную роль за счет весьма быстрого размножения, т.е. весьма быстрого круговорота веществ, связанного с этим размножением.
Количество солнечной энергии видимой части спектра, достигающей земной поверхности и используемой организмами, составляет около 55 ккал/см2 год. Способность накапливать энергию солнечного света в органическом веществе называется продуктивностью живых организмов. Различают валовую первичную продукцию (общее количество органического вещества или связанной в нем энергии, определяется на 1 м2 в год) – это в основном продукция хлорофиллоносных растений, при хемосинтезе бактерий образуется гораздо меньше; чистую первичную продукцию (количество органического вещества или связанной в нем энергии за вычетом расходов на дыхание); вторичную продукцию (продукция организмов, потребляющих или перерабатывающих биомассу, полученную в результате первичной продукции) – это продукция животных и сапробов (потребителей мертвого органического вещества).
Биогенное происхождение имеет большая часть кислорода атмосферы, углекислого газа. Живые организмы влияют на солевой состав гидросферы и являются ведущими агентами почвообразования. По словам В. И. Вернадского, “на земной поверхности нет химической силы, более постоянно действующей, а потому и более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом”.
В настоящее время влияние человека на природу достигло такой степени, что ему приходится соизмерять свою деятельность с возможностями природы. В. И. Вернадским разработано целое учение о ноосфере, представляющей собой сферу взаимодействия природы и человека.
9. Экологические области океана
В океане выделяется несколько типов областей с различными экологическими условиями. Прежде всего различают толщу воды – пелагиаль, область распространения пелагических животных, и бенталь, заселенную бентосом, т. е. организмами, обитающими на грунте или в толще грунта.
Бенталь подразделяют на супралитораль, литораль, сублитораль, батиаль и абиссаль. Супралитораль – побережье океана, расположенное выше уровня самого высокого прилива, но затопляемое при сильных штормах. Здесь обитают как наземные, так и морские организмы.
Литораль – прибрежная зона морского дна, покрытая водой только при приливах. Значительное число видов вне этой области существовать не могут. Только на литорали встречаются вечнозеленые тропические кустарники мангры. В приполярных странах из-за воздействия льда литораль крайне обеднена.
Сублитораль – прибрежная часть океана, непосредственно примыкающая к литорали, простирающаяся в глубину до нижней границы распространения донной растительности (около 200 м). С сублиторалью связаны богатые промыслы беспозвоночных.
Мелководья океана, располагающиеся главным образом в пределах шельфа, называются неретической областью. Для нее характерны большая подвижность воды, значительное проникновение света, большое разнообразие растительного и животного мира. Эта область тесно связана с сушей: обильное органическое вещество доставляется реками, улучшая трофические условия, ряд видов использует и водную, и наземную среду обитания. Здесь встречаются разнообразные водоросли, цветковые растения (руппии, взморниковые). В зарослях, прикрепленных к берегу находят убежище многие животные. На и в мягком грунте обитают разнообразные черви, крабы, морские звезды, моллюски. На скальном грунте встречаются сверлящие животные, животные, прикрепляющиеся к грунту с помощью раковины (усоногие раки, моллюски). Исключительным разнообразием и богатством жизни характеризуются коралловые рифы.
Батиаль располагается на материковых склонах на глубине до 2000 м и характеризуется слабой освещенностью или отсутствием освещенности, колебания температуры и солености незначительны. В покрове дна преобладают илы.
Абиссаль – область наибольших глубин, соответствует ложу Мирового океана. Абиссаль характеризуется чрезвычайно медленным передвижением воды от полюсов к экватору, постоянно низкими температурами воды (около 0°), практически полным отсутствием света. Здесь полностью отсутствуют зеленые растения, накапливаются преимущественно органогенные илы.
Пелагиаль подразделяют на эпипелагиаль – освещенные слои воды над неретической областью и глубоководную пелагиаль, которая, в свою очередь, состоит из батиопелагиали и абиссопелагиали. В пелагиали обитают представители четырех групп организмов: нектона, планктона, плейстона и нейстона (две последние – растения и животные, обитающие на поверхности воды).
Планктон – организмы, обитающие в толще воды и переносимые морскими течениями и волнами, т. к. сами они могут передвигаться лишь весьма медленно. Планктон, как и нектон, имеет специальные приспособления, препятствующие погружению в воду (выработка веществ, имеющих малый удельный вес – жиры и др., насыщение тканей и полостей тела водой). Фитопланктон представляет собой главным образом одноклеточные организмы, имеющие хлорофилл и способные к ассимиляции. Они являются основой жизни океана, источником первичной продукции. Зоопланктон представлен микроскопическими и более крупными организмами – медузами, сифонофорами и даже хордовыми.
Недостаток света становится определяющим для жизни на глубине более 550 м. Здесь полностью отсутствуют продуценты, а, следовательно, и консументы 1-го порядка. Велика роль редуцентов-сапрофагов, потребителей мертвого органического вещества. Общая биомасса уменьшается с глубиной. Из-за почти полного отсутствия движения воды, дно океана покрыто мощным слоем или вулканического или биогенного происхождения. В ил погружаются многие обитатели дна.
Своеобразие обитателей абиссали связано, в первую очередь, с отсутствием света. Большинство обитателей абиссали утратили способность видеть и имеют высоко развитые органы осязания, некоторые способны сами вырабатывать свет.
10. Живое вещество биосферы и его химический состав
Живые организмы избирательно накопляют некоторые химические вещества. Поэтому содержание различных элементов неодинаково в организмах, земной коре и в почвах. Содержание кислорода (70,0%) и углерода (18,0%) в биосфере заметно выше по сравнению с земной корой, а содержание кремния, алюминия, железа – ниже, содержание фосфора и серы – примерно на одинаковом уровне.
Однако организмы различны по своему химическому составу, который может сильно отличаться от среднего. Некоторые виды растений способны к накоплению тех или иных элементов (водоросли – йод и т. д.). Это проявление участия организмов в геохимических процессах. Различный химический состав могут иметь даже разные особи одного вида.
Живое вещество распределено по планете неравномерно. А. П. Виноградов подразделял сушу на биогеохимические зоны (отличаются по содержанию химических элементов и по преобладающим биологическим реакциям организмов на изменчивость геохимических особенностей среды) и биогеохимические провинции (зональные и азональные). Азональные биогеохимические провинции располагаются над рудными телами, в районах вулканизма и бессточных впадинах. В них наблюдаются избыточные концентрации определенных элементов.
Круговорот живого вещества происходит через три основные звена: продуцентов, консументов и редуцентов. Продуценты – фотосинтезирующие зеленые растения и хемосинтезирующие бактерии, т.е. организмы, дающие первичную продукцию; консументы – потребляющие первичную или вторичную, т.е. потребляющие готовое органическое вещество и переводящее его в другие формы органического вещества – животные, паразитические растения; редуценты (деструкторы) – организмы, разлагающие мертвые органические вещества до минеральных веществ – многие бактерии, грибы, простейшие животные.
Консументы делятся на консументов первого порядка – фитофагов, консументов второго порядка – зоофагов и консументов третьего порядка – зоофагов. Встречаются также эврифаги, т.е. всеядные консументы. Многие животные играют также роль редуцентов.
11. Реликт, реликтовый ареал
Реликт – вид, находящийся в противоречии с современными условиями существования. Реликты – это как бы остатки прежних эпох. Реликтовым вид может быть лишь на части своего ареала (например, ель в дубравах). Возникновение реликтов может быть обусловлено разными причинами.
Реликты часто являются свидетелями более широкого распространения той или иной растительной формы, в таком случае они называются формационными.
Реликты эдафические (геоморфологические) являются свидетелями иного, чем в настоящее время распределения суши и моря, песков и грунтов тяжелого механического состава и т. д.
Реликты климатические различаются по тем эпохам, свидетелями климатических условий которых они были. Наиболее древними климатическими реликтами сейчас являются реликты третичного периода (неогена), например, леса Западного Закавказья. Также известно много реликтов ледникового периода, оттесненные ледником далеко на юг и там сохранившиеся (например, тундровые растения в долине р. Чусовой). Третью группу климатических реликтов составляют виды, связанные с последниковым пустынно-степным периодом (например, ксерофильные степные виды в Магаданской области).
Реликтами являются большинство палеоэндемов. Таким образом, реликтовый характер вид приобретает там, где благодаря каким-либо местным особенностям условия существования сходны с характерными в настоящее время для основной площади его ареала (например, склоны северной экспозиции в тайге близки к тундровым условиям и т. п.). Поэтому существуют территории, изобилующие реликтами, и территории, на которых они почти отсутствуют.
12. Понятие “климакс”. Представление об устойчивости биоценозов
На последней стадии сукцессии (см. №19) образуется сложившееся общество. При достаточно долгой совместной жизни на одном и том же участке возрастает экологическая дифференциация. Под пологом светолюбивых растений появляются тенелюбивые, на участок проникают консументы разных порядков; трофические цепи и консорции усложняются. Видовой состав сообщества становится более постоянным, т. е. проникновение на участок новых видов заметно замедляется.
Любые изменения имеют временной предел. Понятие устойчивости биоценоза ввел ученый Клеменс. Биоценозы, характеризующиеся определенной устойчивостью, постоянным видовым составом, хорошо отработанной структурой, находятся в устойчивом равновесии с окружающей средой, называются климаксами. Климакс – открытая система, находящаяся в равновесном состоянии, компоненты которой меняются только в связи с особенностями прохождения жизненного цикла организмов, колебаниями природных режимов и местными локальными нарушениями.
Таким образом, при достижении стадии климакса сообщество не прекращает свое развитие. Развитие сообщества всегда имеет место, поскольку оно происходит не только под влиянием несоответствия общества внешним условиям, но и в результате его самодвижения, без всякого изменения внешних условий.
Климаксовый тип биоценоза – результат длительной сопряженной эволюции всех компонентов растительного и животного мира в глубоком взаимодействии с условиями среды и под контролем климатических условий. Время достижения климакса – сотни лет (до тысячи). Чем жестче природные условия, чем больше видовой состав, тем больше время достижения климакса.
13. Понятия фитоценоз, биоценоз, животное население. Основные подходы к их определению.
Понятия “растительное” и животное население” определенного района характеризуют прежде всего набор растительных (животных) сообществ – фито- и зооценозов. Понятия “флора” и “фауна” характеризуют видовой состав района. Флористическое и фаунистическое разнообразие, таким образом, опредляется числом видов, родов и других систематических категорий.
Постоянно развивающаяся совокупность растений и животных, населяющих участок среды обитания с более или менее однородными условиями существования (биотоп), образовавшаяся естественно или под влиянием деятельности человека, непрерывно развивающаяся и характеризующаяся определенным видовым составом организмов, определенными взаимоотношениями между членами биоценоза и между биоценозом и средой обитания называется биоценозом. Совокупность растений биоценоза называется фитоценозом.
Растения и животные образуют на поверхности Земли сообщества. Сообщество – это общее понятие, прилагаемое к совокупности взаимодействующих живых существ любого ранга. Наименьшей единицей, в которой может быть применен термин “сообщество” является биоценоз.
14. Ценотическая значимость видов. Эдификаторы, соэдификаторы, ассектаторы
Не все виды играют в биоценозе одинаковую роль. Обычно среди растений различают компоненты-многолетники, постоянно присутствующие в биоценозе, и ингредиенты-однолетники, количество которых может резко колебаться год от года в зависимости от условий погоды.
Среди организмов, образующих биоценоз, различают доминанты, т.е. виды преобладающие по численности. Разным систематическим группам организмов, образующих биоценоз, свойственны свои доминанты. Кроме доминантов выделяют также субдоминанты, второстепенные и третьестепенные виды. Критерии разделения на эти группы как правило численные: число особей, биомасса, проективный покров (у растений), обилие. Некоторые ученые выделяют доминанты-растения только для главного яруса, считая господствующие растений второстепенных ярусов субдоминантами. Часто выделяют доминанты групп ярусов и даже отдельных ярусов.
По количеству доминант биоценозы бывают моно-, олиго- и полидоминантные. Часто происходит смена доминант от года к году, вызванная колебаниями параметров внешней среды (“защитная реакция биоценоза”).
Ценотип – группы видов в составе одного биоценоза со сходной ценотической значимостью, близкие сходным влиянием на организацию фитоценоза (биоценоза).
Существуют два основных ценотипа: эдификаторы и ассектаторы. Эдификаторы – доминантные виды, определяющие все основные особенности биоценоза, они обладают наибольшей средообразующей ролью. Эдификаторы таким образом трансформируют окружающую среду, что остальные виды должны подстраиваться. Ассектаторы – постоянные, но не господствующие виды растений в растительном сообществе.
15. Наиболее общие закономерности сукцессий. Смена жизненных форм, видового разнообразия, изменение биомассы.
Флуктуации затрагивают только виды одного сообщества, однако по разным причинам одни сообщества могут сменяться другими. Этот процесс может проходить непоследовательно или последовательно. В последнем случае он называется сукцессией. Сукцессия – такая смена, при которой одно сообщество сменяется другим и которая протекает последовательно, без возврата к прежнему состоянию.
Если сообщество начинает развиваться на участке, полностью лишенном жизни (обсохшие днища морей и озер, песчаные наносы, лавовые потоки, обнажившиеся скалы и осыпи), сукцессия называется первичной.
Вторичная сукцессия – смена одного сообщества другим в результате изменений условий существования или значительных нарушениях состава сообщества.
Вторичные сукцессии могут происходить путем постепенного изменения сообщества в результате изменения им среды обитания или под влиянием внешних воздействий, не связанных с эти логическим развитием. Последний тип смен называется стихийным или внезапным. Внезапные смены могут быть: климатогенными, эдафтогенными, биогенными, пирогенными и антропогенными. Любая смена, начавшаяся под влиянием внешнего по отношению к сообществу фактора, постепенно переходит в эндодинамическую смену, определяемую все в большей степени воздействием сообщества на среду его обитания.
Примером постепенной смены сообществ может служить смена степного сообщества лесным. В северных степях травянистая растительность, постепенно изменяя химизм почвы, создает условия, более благоприятные для существования кустарниковых растений (степная вишня, степной миндаль), чем для трав. В результате травянистые степи сменяются кустарниковыми, которые привлекают таких потребителей плодов кустарников, как обыкновенный хомяк, серая куропатка и др. Среди кустарниковой степи появляются проростки деревьев, и постепенно кустарниковая степь сменяется лесным сообществом с березой и осиной, с грездящимися на деревьях птицами (грач, кобчик), с лесными видами трав и насекомых.
Внезапные смены происходят под влиянием катастрофических воздействий самого различного характера. Каждая такая смена начинается гибелью видов, характерных для данного сообщества, затем происходит постепенно восстановление этого сообщества. Любая сукцессия, вызванная катастрофической причиной, состоит из нескольких быстро сменяющих друг друга начальных стадий после произошедшей катастрофы, а затем значительно более медленное длительное восстановление исходного сообщества.
Общие закономерности:
1. Смена видов различных жизненных стратегий. Виды делятся на раннесукцессионные (эксплеренты и др.) – много семян, высокая скорость роста, светолюбивые, относительно небольшая продолжительность жизни - и позднесукцессионные (эпителенты и др.) – большая продолжительность жизни, тенелюбивые, мало семян, семена не летучие.
2. Изменение числа видов.
3. Изменение соотношения надземной и подземной биомассы (увеличивается доля надзменой), изменение величин самой биомассы (обычно возрастает). Обычно возрастает стабилизирующая роль растительности.
4. Постепенное снижение скорости сукцессии.
16. Зональное распределение живого покрова суши. Важнейшие градиенты среды
Распределение по земному шару биоценозов подчиняется закономерностям разного ранга. Оно зависит от изменения климатической обстановки (соотношение тепла и влаги) с севера на юг – эти объясняется существование природных зон, от окраин материка вглубь – за счет этого возникают провинциальные особенности в пределах зон, а также от сочетания горных пород, подстилающих почвы, определяющих сочетание биоценозов в каждом конкретном участке.
Зональное распределение экосистем подчиняется основному географическому закону – закону географической зональности. Для выявления закономерностей распределения экосистем на земной поверхности в связи с гидротермическими условиями (основные градиенты среды) используется несколько показателей. Наиболее распространен радиационный индекс сухости, предложенный М. И. Будыко:
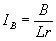 ,
,
где B – годовой радиационный баланс, r – годовая сумма осадков, L – скрытая теплота парообразования. Значениям IB от 1/3 до 1 соответствует распространение лесов (лесная зона), от 1 до 2-х – степная зона, от 2 до 3-х – пустыни и полупустыни. Сочетание показателей В и IB характеризует более точное содержание, т. е. по значениям радиационного баланса можно разделять леса на тропические и умеренного пояса, степи – на степи умеренного пояса и жаркие степи – саванны и т. д.
Еще одни показатель, широко используемый для уточнения зонального распределения экосистем – гидротермический коэффициент (ГТК) Г. Т. Селяниновой:
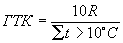 ,
,
где R – количество осадков за период, когда t > 10°C. Северная граница степей совпадает со значением ГТК = 1, пустынной – 0,5, широколиственных лесов – 1,5.
Основными показателями, интегрально отражающими структуру и функционирование экосистем и непосредственно зависящими от гидротермических условий, являются запасы фитомассы.
17. Сообщества организмов океана
См. вопрос №9.
18. Флуктуации биоценозов
Погодичная (разнодинамичная) динамика, или флуктуация, характеризуется ненаправленностью, обратимостью происходящих изменений, т. е. возможностью возврата сообщества к состоянию, близкому к наблюдавшемуся несколько лет назад, отсутствием резкой смены флористического и фаунистического состава. При малом числе трофических уровней и незначительном разнообразии добычи, доступной каждому виду (например, в тундре) любое значительное колебание численности одних животных оказывает воздействие на обилие связанных с ними трофически других. В таких случаях происходят большие флуктуации численности животных всех трофических уровней.
Хорошим примером флуктуаций являются изменения, происходящие в широколиственном лесу, связанные с изменениями урожайности плодов деревьев, которая различна в различные годы, причем хорошие урожаи случаются раз в 2-4 года. С отставанием на год от высокого урожая семян наблюдается повышение численности грызунов и птиц, их потребляющих, а с отставанием еще на год – повышение численности иксодовых клещей, личинки которых питаются преимущественно кровью грызунов. На год высокой численности зверьков или на следующий год приходится пониженный урожай семян, поэтому рост численности животных прекращается, а затем численность начинает падать. На этот период приходится подъем урожайности семян и все начинается сначала.
19. Первичные сукцессии. Стадии формирования биоценоза
Сукцессия – такая смена, при которой одно сообщество сменяется другим и которая протекает последовательно, без возврата к прежнему состоянию.
Если сообщество начинает развиваться на участке, полностью лишенном жизни (обсохшие днища морей и озер, песчаные наносы, лавовые потоки, обнажившиеся скалы и осыпи), сукцессия называется первичной. Первичная сукцессия проходит три последовательные стадии. Заселение лишенной жизни поверхности определяется случайностями заноса живых существ и свойствами субстрата.
Сначала появляются случайно занесенные сюда растения – пионерная группировка. Для многих условия оказываются неподходящими, они не достигают зрелости и не приносят потомства. Как правило, успешно развиваются на таком участке лишь немногие виды. Пионерная группировка характеризуется неопределенностью состава (одни виды появляются, другие исчезают), разобщенностью отдельных экземпляров растений, которые еще не могут оказывать влияние друг на друга, но уже оказывают в той или иной степени влияние на окружающую среду. Таким образом, сообщество еще не является замкнутым (т. е. возможно вселение новых видов) и не является сомкнутым (надземные и тем более подземные части растений не смыкаются друг с другом).
На втором этапе формирования биоценоза образуется группировка. Растения начинают возобновляться семенами или вегетативным путем, животные размножаются. Темпы заселения участка возрастают, а т. к. источником молодых экземпляров становятся преимущественно особи, уже обитающие на этом участке, то сложение растительного покрова становится зарослевым; животное население приобретает пятнистый характер (чередование скоплений и разрежений). Состав такого сообщества не вполне устойчив, довольно легко происходит внедрение новых видов, виды оказывают влияние друг на друга только по границам пятен и групп, структура сообщества еще не устоявшаяся, но довольно пестрая. Значительную роль начинает играть конкуренция.
Наконец образуется сложившееся общество. При достаточно долгой совместной жизни на одном и том же участке возрастает экологическая дифференциация. Под пологом светолюбивых растений появляются тенелюбивые, на участок проникают консументы разных порядков; трофические цепи и консорции усложняются. Видовой состав сообщества становится более постоянным, т. е. проникновение на участок новых видов заметно замедляется. По мере развития сообщество приходит во все большее соответствие с условиями среды. Сообщество, соответствующее тем условиям, в которых оно существует, называется климаксом.
21. Круговорот кислорода
Первоначальным источником всех процессов, протекающих на Земле, является Солнце. Основной процесс, в результате которого образуется свободный кислород, а также синтезируются органические вещества (углеводы), является фотосинтез.
Фотосинтез – у зеленых растений процесс преобразования света в химическую энергию органических соединений, синтезируемых из диоксида углерода и воды. Фотосинтез представляет собой цепь окислительно-восстановительных реакций, совокупность которых принято разделять на две фазы – темновую и световую. КПД фотосинтеза очень низкое (менее 1%), это связано с малым содержанием углекислого газа в атмосфере и гидросфере. Суммарное уравнение процесса фотосинтеза выглядит так:
6СО2 + 6Н2О 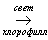 C6H12O6 + 6O2
C6H12O6 + 6O2
Фотосинтез – единственный биологический процесс, который идет с увеличением свободной энергии и прямо или косвенно обеспечивает доступной химической энергией все земные организмы (кроме хемосинтезирующих, получающих энергию за счет окисления неорганических веществ – аммиака, водорода, соединений серы, оксида железа II). Процессу фотосинтеза обязано своим существованием все живое на Земле. Он является практически единственным источником свободного кислорода на Земле.
Гораздо меньшее количество кислорода образуется при фотолизе – разложении воды на кислород и водород под действием света.
Дыхание – совокупность процессов поступления в организм кислорода и удаления углекислого газа (внешнее дыхание), а также использование кислорода клетками и тканями для окисления органических веществ с освобождением энергии, необходимой для их жизнедеятельности (клеточное или тканевое дыхание). У одноклеточных животных и низших растений обмен газов при дыхании происходит путем диффузии через поверхность клетки, у высших растений – через межклетники, пронизывающие все тело. У многоклеточных животных внешнее дыхание осуществляется специальными органами дыхания, а тканевое – обеспечивается кровью.
22. Пресные водоемы как среда жизни
Одним из уникальных свойств пресной воды является то, что она обладает наибольшей плотностью при 4°С, при более высоких или низких температурах она расширяется и становится легче. Поэтому лед образуется только в приповерхностном слое водоема. Различают стоячие и проточные внутренние водоемы. Эти типы водоемов связаны переходными формами – старицы, проточные озера, временные водотоки.
Проточные водоемы обычно имеют пресную воду. Соленость стоячих водоемов существенно различается по количеству и составу солей. Содержание карбоната кальция называется жесткостью воды, оно является важным регулирующим фактором жизни в пресных водоемах. Водоемы с самой жесткой водой являются пресными.
У обитателей пресных вод жидкости их тел гипертоничны, т. е. концентрация солей в них выше, чем в воде. По законам осмоса вода стремится проникнуть в их тела. Поэтому существуют водозащитные или водоудаляющие механизмы.
Во внутренних водоемах резко варьирует содержание органических веществ и связанное с ним содержание кислорода. Содержание кислорода выше всего в водах быстротекущих рек, также велико содержание органики в болотах. Излишнее количество органики может привести к цветению водоема и гибели организмов.
Температурный режим характеризуется весенним и осенним перемешиванием, летним прямым распределением и зимним обратным (вспомним гидрологию). Слой температурного скачка между эпилимнионом и гиполимнионом называется термоклином. Организмы, способные жить при определенной маломеняющейся температуре, называются стенотермными, при различных температурах – эвритермными.
Движение воды во внутренних водоемах представлено течениями (в проточных водоемах) и волнениями (в крупных озерах). Различают реофильные (обитающие в быстротекучих реках и ручьях) и лимнофильные (обитающие в стоячих водах) организмы.
По условиям существования для растений и животных стоячие водоемы могут быть поделены на олигтрофные (большие глубины, плохо развита литоральная растительность, малая первичная продукция), мезотрофные и эвтрофные (все наоборот).
Особыми условиями характеризуются подземные водоемы (отсутствие света, фотосинтеза, волнений, слепые животные), водохранилища (обилие ила, следовательно, затрудненное икрометание; заболоченные берега).
Внутренние водоемы тесно связаны с окружающей средой. Огромное значение имеет сток с суши минеральных и органических веществ. Внутренние водоемы влияют на климат и гидрологический режим окружающей территории. Многие виды растений и животных связывают водную среду обитания с наземной (тростник, камыш, бобр, выдра). Обилие во внутренних водоемах космополитных и других широко распространенных видов связано с их способностью перемещаться из водоема в водоем, широкой экологической амлитудой.
23. Подходы к выделению жизненных форм. Жизненные формы животных
Для животных, как и для растений, важная система жизненных форм, основанная на отношении к климатическим особенностям. Именно с этим фактором связана классификация жизненных форм растений К. Раункиера. Но для животных имеют существенное значение общие особенности среды обитания, способ передвижения в этой среде, а также характер их пищи. Непосредственное же воздействие климата уменьшается для гомотермных животных потому, что их физиологические особенности способствуют выработке некоторой независимости от его влияния.
Д. Н. Кашкаров разработал несколько классификаций жизненных форм животных. Первая из них имеет следующий вид:
1. Холоднокровные (пойкилотермные животные): 1) деятельные весь год; 2) недеятельные часть года: а) летоспящие, б) зимоспящие.
2. Теплокровные (гомотермные): А. Оседлые. 1) деятельные весь год; 2) недеятельные часть года: а) летоспящие, б) зимоспящие. Б. Сезонные. 1) гнездящиеся, 2) зимние, 3) летние, 4) пролетные.
По способам передвижения в различной среде обитания Д. Н. Кашкаров выделяет такие жизненные формы:
1. Плавающие. 1) чисто водные: а) нектон, б) планктон, в) бентос; 2) полуводные: а) ныряющие, б) неныряющие, в) лишь добывающие из воды пищу.
2. Роющие. 1) абсолютные землерои (всю жизнь проводят под землей), 2) относительные землерои (выходят на поверхность).
3. Наземные. 1) не делающие нор: а) бегающие, б) прыгающие, в) ползающие; 2) делающие норы: а) бегающие, б) прыгающие, в) ползающие; 3) животные скал.
4. Древесные, лазающие: а) не сходящие с деревьев, б) лишь лазающие на деревья.
5. Воздушные формы: а) добывающие пищу в воздухе, б) высматривающие пищу с воздуха.
Если к этим классификациям добавить еще две: по отношению к влажности воздуха и почв (влаголюбивые и сухолюбивые) и по месту размножения (под землей, на поверхности земли, в ярусе трав, на ветвях деревьев, в трещинах и дуплах), то эта совокупность классификаций определит основные признаки жизненных форм животных.
Конкретных примеров отнесения животных к разным группам в различных системах жизненных форм Д. Н. Кашкаров не приводит.
Для организмов имеет существенное значение не только приспособение к условиям жизни, но и способность к расселению, позволяющая менять место своего обитания. К перемещениям организмов в пространстве относятся две группы явлений: периодические перемещения – миграции и непериодические – инвазии (нашествия). Если непериодическое перемещение заканчивается вымиранием видов, то оно не является инвазией. Миграции чаще всего связаны с особенностями климата. Инвазии обычно обусловлены неблагоприятной кормовой обстановкой в местах существования данного вида.
Совершают миграции или инвазии группы совместно обитающих особей, относящихся к тому или иному виду - популяции. Популяция - часть населения вида, занимающая определенную площадь, в пределах которой происходит обмен генетической информацией. Различают наиболее крупные климатические (у животных – географические) популяции, более мелкие эдафические (у животных – экологические) и ценотические, фитоценотические (у животных – элементарные) популяции. Популяции различаются по образу жизни, основным кормам, численности, степени процветания, степени соответствия условиям обитания.
24. Межвидовые взаимоотношения в биоценозе. Принцип конкурентного исключения и дифференциации экологических ниш.
Организмы, живущие в биоценозе вступают в следующие биотические отношения:
| тип отношений | для А | для Б | пример, объяснение |
| мутуализм | + | + | рак-отшельник (передвигает раковину в места с лучшими условиями питания) и актиния (дает доп. защиту от врагов) |
| комменсализм | + | 0 | рыбы-прилипалы (двигаются, питаются остатками пищи хозяев) и рыбы-хозяева |
| хищничество, паразитизм | + | - | консументы и продуценты, либо консументы более низкого порядка |
| нейтрализм | 0 | 0 | без комментариев |
| антибиоз | 0 | - | выделение токсичных веществ, не вредных для выделяющего организма |
| конкуренция | - | - | межвидовая и внутривидовая |
Функциональная направленность таких отношений может быть различной. Наиболее широко распространены трофические (пищевые) отношения. Именно ими определяются трофические уровни: продуценты, редуценты, консументы трех порядков. По набору кормов различают эврифагов (широкий круг кормов) и стенофагов (узкий круг кормов). В пределах этих групп выделяют формы животных: растительноядные, животноядные и со смешанным питанием. Каждая из этих групп делится на семеноядов (потребители зеленых кормов, древесины, коры растений; мяса и других тканей живых животных, трупов, грибов) и копрофагов (потребители экскрементов, отмерших растительных тканей и т. п.). Каждое последующее звено пищевой цепи дает меньшую массу, чем предыдущее, поэтому любая пищевая цепь характеризуется пирамидой биомасс.
Топические отношения связаны с представлением одними видами места поселения другим. Чаще всего в качестве хозяев “квартиры” выступают растения, однако известны, например, водоросли, живущие в шерсти ленивца. Известно значительно число животных нидиколов – обитателей нор других животных. Постоянно пребывание на определенном субстрате – причина возникновения окраски и формы, получивших название покровительственных.
Форические отношения – отношения между видами, когда одни виды способствуют перемещению с места на место других видов. Фабрические отношения – один вид животного использует части другого животного или растения при строительстве гнезд, подстилки в норах и т. п.
Для понимания роли тех или иных организмов в жизни сообщества имеет существенное значение оценка их численности. Можно подразделять виды по степени их доминирования, имеются и другие способы оценки численности (обилия) вида. Различают абсолютную (в цифрах) и относительную (в баллах или %) оценку обилия. При цифровой оценке определяются количество особей на единицу площади, объем вида на единицу объема сообщества, массу вида на единицу площади, объема или в пересчете на число особей.
Методы балльной оценки численности дают значительно менее точные результаты, но они и менее трудоемки. Для растений обычно применяют пятибалльную шкалу О. Дроде. Относительные оценки численности у растений основываются обычно на проценте площади, занятой проекциями наземных частей от общей площади; наименьших расстояний между растениями; сумме поперечных сечений или площадей основания стволов. Для животных используется, как правило, трехбалльная шкала: вид многочислен, обычен, редок. В качестве критериев косвенной оценки могут служить: количество нор, подсчет по голосам, количество следов на единицу площади, число кормовых столиков и пр.
В биоценозе можно выделить структурно-функциональные единицы – консорции. Под консорцией понимают совокупность самостоятельно существующего (т.е. не нуждающегося в опоре и не поселяющегося на другом виде) зеленого растения – центра консорции – со всеми организмами, связанными с ним трофическими, топичесими, форическими и фабрическими связями, т. е. использующими его в процессе жизнедеятельности - консортами. Консорции тесно переплетаются друг с другом, поскольку одни и те же виды консортов могут быть связаны с различными центрами консорций.
Конкурентные взаимоотношения являются наиболее всеохватывающими в природе. Конкуренция может быть межвидовой и видовой. Межвидовая конкуренция имеет место в том случае, когда особи разных видов претендуют на одни и те же ресурсы или подвергаются одной и той же опасности. Внутривидовая борьба происходит между особями одного и того же вида. В результате конкуренции происходит дифференциация организмов по экологическим нишам, некоторая часть особей отмирает.
25. Представление о континууме. Границы биоценозов
Большинство ученых придерживается представления о постепенности, континуальности растительного покрова (Л. Г. Раменский и др.). При этом речь может идти о континууме топографическом, темпоральном и таксономическом (постепенные переходы между таксономическими категориями). Животные в отличие от растений, менее тесно связаны с сообществами. Поэтому при решении вопроса о резкости или континуальности границ основное значение имеют неподвижные, привязанные к месту растения.
К вопросу о резкости границ между биоценозами следует подойти с таких позиций:
1. Плавность границ сообществ зависит от масштаба их изображения и изучения.
2. Чем ниже таксономическая категория, тем более плавными являются ее границы.
3. Одни из сообществ обладают более резкими, другие – более плавными границами; более резкими границами характеризуются сообщества, эдификаторы которых оказывают более мощное преобразовательное воздействие на среду.
4. Как правило, условия существования сообществ сменяют я более постепенно, чем сами сообщества.
5. Возникающая в процессе развития сомкнутости сообществ сопряженность между образующими их видами способствует большей резкости границ между сообществами.
Таким образом, хотя непрерывность биоценотического покрова нашей планеты не вызывает сомнения, однако в нем могут быть выделены участки, однородные по видовому составу, характеру динамики и структурным особенностям. Они отделены от соседних участков то резкими, то плавными постепенными переходами, однако и в последнем случае ширина переходной полосы много меньше, чем ширина участков, которые она связывает.
26. Биоценоз и условия местообитания. Средообразующая роль живых организмов
Растения и животные образуют на поверхности Земли сообщества. Сообщество – это общее понятие, прилагаемое к совокупности взаимодействующих живых существ любого ранга. Наименьшей единицей, в которой может быть применен термин “сообщество” является биоценоз. Границы биоценоза и связанного с ним биотопа обычно определяют по изменениям растительного покрова.
Любой биоценоз изменяет среду обитания, с которой он связан. Сообщество характеризуется своей внутренней средой, отличающейся от среды, не преобразованной сообществом.
Особенно заметны эти изменения в сомкнутых, в первую очередь лесных биоценозах, менее выражены они в биоценозах открытых. Так, в лесном биоценозе снижены скорость ветра, амплитуды температур, средняя температура, количество света, проникающего к поверхности почвы, увеличена влажность воздуха, формируются почвы определенного состава.
Таким образом, организмы, обитающие в пределах того или иного биоценоза, находятся под влиянием условий среды, видоизмененной компонентами этого биоценоза. В любом биоценозе организмы образуют его неподвижный каркас, а подвижные животные проводят время в различных частях этого биоценоза, время от времени покидая его и вновь возвращаясь, поскольку выполнение различных функций (размножение, питание, выращивание потомства) может быть связано с различными биоценозами.
Внутренняя среда сообщества далеко не однородна. Каждый вид, каждая жизненная форма, образующие сообщество, находят в нем для себя наиболее благоприятные условия существования. Нередко для выполнения разных функций одним и тем же видам необходимы различные условия. Таким образом, сообщество представляет сочетание разнообразных в экологическом отношении групп.
Очень существенное значение имеет характер пребывания видов в биоценозе. Растения, как правило, неподвижны. Группы растений, образующие сообщество, различаются тем, какую долю своих органов они теряют в неблагоприятные периоды годы. Животные менее тесно связаны с сообществом. Различают изменения характера пребывания животных в сообществе по фазам метаморфоза, времени суток и сезонам. Соотношения между животными и растениями различных групп в разных географических зонах, конечно же, неодинаково.
По привязанности к сообществу все организмы могут быть поделены на две основные группы: сесильные (постоянные) – растения, грибы, бентосные формы в водных сообществах, и вагильные (непостоянные) – большая часть животных и неприкрепленные водные растения.
27. Биоценоз, биогеоценоз, экосистема
Совокупность условий, необходимых для существования того или иного вида, называется стацией (для растений – местообитание). Участок земной поверхности, характеризующийся определенными абиотическими условиями, называется биотопом. Постоянно развивающаяся совокупность растений и животных, населяющих участок среды обитания с более или менее однородными условиями существования (биотоп), образовавшаяся естественно или под влиянием деятельности человека, непрерывно развивающаяся и характеризующаяся определенным видовым составом организмов, определенными взаимоотношениями между членами биоценоза и между биоценозом и средой обитания называется биоценозом.
Биоценоз в совокупности с биотопом образует биогеоценоз (термин введен В. Н. Сукачевым). Биогеоценоз представляет собой совокупность абиотических и биотических компонентов природы. Биогеоценоз – самое мелкое из подразделений земной поверхности. Если в пределах какой-либо фации есть жизнь, и ни одна ее часть не изменена человеком, то она представляет собой биогеоценоз.
В качестве безразмерной категории при подразделении земной поверхности применяется термин “экосистема”. Экосистема – совокупность взаимосвязанных природных компонентов и, в некоторых случаях, человека, способных функционировать как единое целое, продуцируя новое вещество. Это последнее определение было предложено профессором Николаевым В. А. (лектор по ландшафтоведению). Если биогеоценоз является пространственной единицой определенной величины, ограничиваемой от соседних биогеоценозов по фитоценозу, то экосистема – единица функциональная, которую не обязательно пространственно отграничивать от других экосистем. Экосистема, в отличие от биогеоценоза, может быть полностью искусственной, например, аквариум (тоже, между прочим, БГ).
28. Жизненные формы растений. Классификация К. Раункиера
Жизненная форма – группа организмов, сходных по облику благодаря жизни в близких или одинаковых условиях, имеющих близкие или одинаковые экологические особенности.
Классификация жизненных форм датского ботаника К. Раункиера построена с учетом отрицательных факторов неблагоприятного периода и защитной роли снежного покрова. При выделении жизненных форм учитываются: положение почек возобновления по отношению к поверхности почвы, а при более дробном делении – степень защищенности почек и наличие или отсутствие листьев в неблагоприятный период года и др. Основные жизненные формы по К. Раункиеру:
1. Фанерофиты - растения, почки и концевые побеги, предназначенные для переживания неблагоприятного периода, расположены на стеблях, живущих несколько, иногда много лет. Фанерофиты подразделяются на 15 подтипов в зависимости от высоты деревьев или кустарников, наличия или отсутствия почечных чешуй, сохранения или опадания листьев в неблагоприятный период. К этому типу также относятся травянистые фанерофиты (тропические растения с травянистыми многолетними стеблями) и эпифиты (часто выделяются в отдельную жизненную форму) – растения, развивающиеся на других растениях.
2. Хамефиты – растения, почки и концевые побеги которых, предназначенные для перенесения неблагоприятного периода, находятся на побегах или частях побегов, лежащих на поверхности или располагающихся настолько близко к ней, что покрываются снегом или мертвыми останками растений, где снега нет. К этому типу относятся полукустарники, многие травы и кустарники с ползучими или приподнимающимися побегами.
3. Гемикриптофиты – растения, побеги которых отмирают в самом начале неблагоприятного периода до уровня почвы и остаются только нижние части растений, защищенные почвой и отмершими листьями. К гемикриптофитам относятся виды, у которых имеются розетки или самые крупные листья располагаются при основании стебля, а также растения, у которых воздушные побеги удалены от основания, а у основания развиты чешуйки. К этому же типу относится малина, у которой развиваемые в первый год жизни побеги являются вегетативными, а генеративные побеги вырастают на боковых ветвях вегетативных побегов.
4. Криптофиты – растения, у которых почки или окончания побегов, предназначенные для перенесения неблагоприятного периода, расположены ниже поверхности почвы или на дне водоемов. К этому типу относятся геофиты (корневищные, клубневые, клубнелуковичные, луковичные, корневые), а также водные растения – гидрофиты, частично или полностью погруженные в воду.
5. Терофиты – переживают неблагоприятный период в виде семян.
По соотношению между числом видов, относящихся к различным жизненным формам, К. Раункиер строил биологические спектры, на которых в процентах показано число видов, относящихся к той или иной жизненной форме.
29. Факторы, обуславливающие верхний предел жизни в горах. Специфика высокогорных биоценозов.
Факторы, определяющие высотную поясность:
1. Изменение температуры. От подножия к вершине горы происходит понижение температуры. Оно составляет 0,5-0,7° на каждые 100 м. Иногда происходит инверсия температуры – повышение температуры при росте абсолютной высоты. Инверсии температуры в горах связаны со стеканием холодного воздуха в долины и котловины.
2. Изменение количества осадков. Как правило, количество осадков с высотой увеличивается, но до определенной высоты, называемой зоной максимальных осадков. Выше количество осадков падает вследствие низких температур и низкой абсолютной влажности.
3. Изменение атмосферного давления. Атмосферное давление воздуха и парциальное давление кислорода с высотой понижаются. На уровне 5500 м атмосферное давление вдвое меньше приземного.
4. Характер горных пород.
5. Экспозиция склонов.
6. Крутизна склонов.
Как правило, в высокогорьях развиты биоценозы, называемые альпийскими и субальпийскими лугами. Также встречаются горная тундра, гольцы и высокогорные пустыни. Для подобных видов растительности характерен короткий вегетационный период, что приводит к выработке особых жизненных форм, близких к жизненным формам обитателей арктической зоны. Так, исключительное господство получают хамефиты, наземные части растений развиты плохо. Низкие температуры воздейха приводят также к развитию таких жизненных форм, как шпалерные формы, растения-подушки и т. п.
Высокогорные биоценозы имеют много общего с биоценозами тундры, однако отличаются большим количеством осадков, отсутствием вечной мерзлоты, особым световым режимом, особым спектральным составом света (меньше доля длинноволнового излучения).
30. Жизненные стратегии видов. Фитоценотипы Л. Г. Раменского
Фитоценотип – группа видов растений в составе одного биоценоза со сходной ценотической значимостью, близких сходным влиянием на организацию фитоценоза.
Жизненная стратегия – совокупность адаптаций, обеспечивающих возможность совместного сообитания и занимания в сообществе определенных позиций. Несколько крупных фитоценотипов по жизненным стратегиям выделил Л. Г. Раменский. Другой биолог Джон Грайм выделил три основных жизненных стратегии для растений:
1. Виоленты (силовики) – растения, обладающие набольшей конкурентоспособностью, максимально использующие ресурсы среды, характеризующиеся высокой энергией жизнедеятельности. Пример: ель. Такой стратегией также обладают крупные животные, для которых характерно малое потомство. Их численность сильно регулируется внешними факторами.
2. Патиенты (терпеливцы) – все адаптации направлены на то, чтобы существовать при недостатке ресурсов.
3. Эксплеренты (выполняющие и заполняющие) – очень высокая энергия роста и размножения. Эти виды ориентированы на захват любых освободившихся участков (где снята конкуренция). Для них характерна быстрая смена поколений. Эти виды обычно занимают местообитания с нестабильными условиями.
31. Соленость как экологический фактор в распространении морских организмов
Для жизни обитателей океана несомненно велико значение солености его вод. Средняя соленость вод Мирового океана составляет 35 промилле. Среднегодовая соленость колеблется от 30 промилле в приполярных областях до 37,9 промилле в области пассатов. В приполярных областях весной при таянии льдов соленость может опускаться до 15 промилле. Окраинные и внутренние моря могут иметь соленость, которая существенно отличается от солености океана.
Падение солености влечет за собой уменьшение разнообразия мира ее обитателей. Так, рифообразующие кораллы не развиваются напротив устьев рек, где вода слишком пресная. Специфические морские виды исчезают в морях с пониженной соленостью. Существуют и эвригалинные виды.
Соленость океанической воды требует определенного осмотического давления внутренних растворов организмов. Многие беспозвоночные животные изотоничны (это связано с тем, что они искони обитают в этой среде). Позвоночные животные имеют ряд приспособлений, обеспечивающих возможность их существования в морской воде. Они, как правило, гипотоничны и гомеоосмотичны (содержание солей понижено, осмотическое давление постоянно). Для поддержания постоянного осмотического давления они пьют морскую воду, выделяя избыток солей при помощи жабр, почек. Некоторые обеспечивают постоянство осмотического давления благодаря непроницаемым покровам из хитина или роговых образований.
Из растворенных солей специфическое значение имеет углекислый кальций (3,6% солей). Эта соль необходима для построения коралловых рифов, раковин моллюсков. Усвоение ее понижается при низкой температуре, поэтому большинство использующих ее организмов обитает в верхних слоях тропических вод.
32. Центры таксономического разнообразия в ареале
Н. И. Вавилов установил, что вид неоднороден на протяжении его ареала, причем эта неоднородность определяется географическими и экологическими особенностями его обитания. Согласно, закону гомологических рядов, согласно которому наследственная изменчивость родственных видов и родов в значительной степени сходна, т. е. амплитуда и характер изменений наследуемых признаков оказываются у близких видов и родов похожими. Этот закон имеет большое практическое значение при поисках сортов культурных растений и пород домашних животных с теми или иными требуемыми признаками.
Первоначальная сплошная область распространения вида (целая популяция) называется центром происхождения вида. Часть ареала, в которой изменчивость данной систематической группы является наибольшей, называется центром многообразия форм. Как правило эти центры совпадают, т. к. в там, где вид существует дольше, он имеет больше возможностей видоизменяться. Однако при значительном изменении условий существования, центр многообразия форм может существенно отличаться от центра происхождения вида. Согласно миграционной теории возникновения ареала, вид расселяется из центра происхождения до тех пор, пока не достигнет пределов, далее которых он распространиться не может.
Вавилов также установил, что существуют центры происхождения культурных растений, которые, как и любые центры возникновения видов характеризуются усиленной изменчивостью, а, следовательно, и сосредоточением большого числа сортов (подвидов). Как правило, здесь наблюдается обилие гетерозиготных форм, т. е. форм, получивших свои гены от разнородных по генетическим особенностям родителей, дающих расщепление признаков при дальнейшем разнмножении, т. е. дающих разнородное по признакам потомство. Обычно такие центры приурочены к горным странам, т. к. именно здесь значительная неоднородность территории вызывает и большое сортовое разнообразие. Именно наличие более или менее значительной амплитуды высот приводит к тому разнообразию условий, которое необходимо для интенсивного формообразования.
33. Биогеографическое районирование Мирового океана
Биогеография океана – наука о распространении и распределении организмов в океане и расчленении океана на биогеографические регионы различного ранга: области, подобласти, провинции. Границы между биогеографическими регионами в океане значительно более постепенны, плавны, чем на суше.
Условия существования в бентали и пелагиали, в неретической и океанической (глубоководной) областях, в освещенной и неосвещенной частях настолько различаются, что следовало бы проводить разделение Мирового океана отдельно для каждой из этих областей жизни. Однако, как правило, необходимое для этого обилие биогеографических данных отсутствует. Обычно в пределах каждой крупной области Мирового океана выделяют одну-две океанические подобласти и часто несколько неретических.
Мировой океан подразделяют на семь биогеографических областей: Арктическую, Бореально-Атлантическую, Бореально-Тихоокеанскую, Тропико-Атлантическую, Тропико-Тихоокеанскую, Нотально-Антарктическую (Субантарктическую) и Антарктическую. Биогеографическое районирование земного шара носит зональный характер.
34. Понятия “флора”, “фауна”. Основные подходы к их изучению
Флора – совокупность видов растений, микроорганизмов и грибов, обитающих в данном районе. Фауна – совокупность видов животных, обитающих в данном районе. Флора и фауна того или иного района составляют в совокупности его биоту.
Понятия “растительное” и животное население” определенного района характеризуют прежде всего набор растительных (животных) сообществ – фито- и зооценозов. Понятия “флора” и “фауна” характеризуют видовой состав района. Флористическое и фаунистическое разнообразие, таким образом, определяется числом видов, родов и других систематических категорий.
Термины “флора” и “фауна” могут быть применены не только ко всей совокупности растений, грибов, микроорганизмов или животных, но и к крупным подразделениям растительного или животного мира (например, флора цветковых растений, флора споровых растений, флора папоротников, микофлора, микрофлора, фауна млекопитающих и т. д.).
Флору и фауну любого региона суши подразделяют на флористические и фаунистические элементы, проводя тем самым флористический и фаунистический анализ. Прежде всего выделяются автохтоны, возникшие в данном месте, и мигранты – относительно недавние переселенцы на данную территорию. При длительном пребывании (давние переселенцы) вида образуются новые виды, являющиеся поэтому автохтонными. Элементы флоры и фауны подразделяют на географические и генетические.
35. Ареал. Ареалы космополитные и эндемичные
Ареал – область распространения той или иной систематической категории. Размеры ареалов могут быть различными. Имеются весьма обширные – космополитные и совсем незначительные – эндемичные ареалы. Размеры ареала зависят от вагильности (подвижности) вида, его экологической валентности и его возраста. Величина ареала прямо пропорциональна экологической амплитуде вида. Сужение наших представлений об объеме вида может привести к уменьшению числа ареалов, которых мы называли космополитными.
Абсолютных космополитов, т. е. распространенных по всей суше (по всему океану) практически не существует. Поэтому обычно под космополитным ареалом понимается обширный ареал, части которого приурочены к значительному кругу местообитаний. Условно космополитами называют виды, занимающие более ? площади суши или более ? акватории Мирового океана. Иногда выделяют полукосмополитов и тропикополитов.
Космополитическими являются некоторые обитатели внутренних водоемов – виды рдестов, ряски; серый дельфин, касатка, черный ворон, скопа, сокол-сапсан. Обширный набор космополитов встречается среди сорных и мусорных растений и сопровождающих человека животных – синантропов (обыкновенный одуванчик, пастушья сумка, серая и черная крысы, домовая мышь).
Под эндемичными видами (эндемами) понимаются виды, ареалы которых не выходят за пределы какой-либо естественной области, ограниченной от соседних. Обычно верхним пределом размеров такой области является биотическая провинция.
Видовой эндемизм особенно характерен для горных вершин, резко отграниченных от равнинных пространств или других горных вершин; островов, ранее составлявших части континента, ныне изолированных водой от остальной суши; участков, отграниченных природными условиями от других сходных участков (например, островные березовые лески полупустынь Казахстана, в которых произрастает киргизская береза).
Эндемы могут иметь различный возраст. Древние эндемы, или палеоэндемы, связаны с филогенетической обособленностью и эколого-географической изолированностью (приурочены горным системам, древним островам). Палеоэндемами также являются виды, сузившие свой ареал в результате изменения обстановки. Молодые эндемы, или неоэндемы, обычно появляются там, где какая-либо более древняя форма в изменившихся условиях обитания дает начало более молодой. Часто это не виды, а подвиды и популяции. Палеоэндемы не имеют близких таксономически видов в других областях, неоэндемы имеют.
36. Вертикальная структура биоты Мирового океана
См. вопрос №9.
37. Сезонная изменчивость биоценозов. Смена аспектов
Для сообществ характерна сезонная смена их облика, так называемая смена аспектов, т. е. в смене сезонных фаз развития, которым свойственна определенная физиономичность (внешний вид).
Сена аспектов связана с изменением состава обитающих в этих сообществах животных и фенологического (сезонного) состояния образующих эти сообщества растений. Разновременное развитие растений в фитоценозе отражает особенности жизнедеятельности растений и их приспособления к изменениям метеорологических условий и фитоклимата.
Сезонная изменчивость позволяет на единице площади существовать большему числу видов. Более ярко смены аспектов выражены в открытых фитоценозах (например, в степях), менее ярки, хотя также хорошо заметны, они в закрытых (например, лесных) фитоценозах. Сообщества могут быть с постоянными аспектами (древесные, кустарниковые) и с меняющимися аспектами в течение вегетационного периода.
Разные виды биоценоза играют разную роль в формировании того или иного аспекта (шкала физиономичности В. В. Алехина). Среди травянистых растений, создающих разные аспекты при массовом их цветении, можно отметить прострел, горицвет весенний, ковыль, незабудку.
38. Биогенный круговорот углерода (С)
Соединения углерода постоянно возникают, изменяются и разлагаются в биосфере. Основной путь углерода – от углекислого газа в живое вещество и обратно – из живого вещества в углекислый газ. При этом часть углерода на долгое время выходит из круговорота, отлагаясь в осадочных породах океана или консервируясь в виде ископаемых горючих веществ органического происхождения - каустобиолитов.
Важнейший процесс, ведущий к образованию органического вещества из неорганического, - фотосинтез; организмы, участвующие в этом процессе называются фитоавторами. Они используют для построения органического вещества энергию солнечных лучей. Хемотрофы используют для построения органического вещества энергию химических реакций и играют гораздо меньшую роль.
Процесс окисления тканей живых существ, при котором выделяется углекислый газ, представляет собой дыхание. Дыхание происходит круглосуточно. Дышат не только ассимилирующие кислород организмы, но и все остальные, за исключением анаэробных. Источником углекислого газа являются и разлагающиеся мертвые органические вещества. Деятельность консументов различных порядков приводит к преобразованию входящих в состав их пищи органических веществ.
Круговорот углерода в океане в значительной степени автономен. Органическое вещество, вырабатываемое фитопланктоном и в значительно меньшем количестве фитобентосом (обитает на мелководье), потребляется зоопланктоном, зообентосом и некотоном. Углекислота, выделяемая ими в процессе дыхания, а также образующаяся при разложении их трупов, растворяется в морской воде.
Существует и обмен углекислотой между атмосферой и океаном, которому благоприятствуют ветер и движение воды. Поэтому количество углекислого газа, растворенного в верхних слоях океанической воды, находится в равновесии с содержанием его в атмосфере. Мировой океан регулирует содержание СО2 в атмосфере путем превращения бикарбонатов в карбонаты (выделяется СО2) при недостатке СО2 в атмосфере и наоборот.
Из огромного запаса углерода, имеющегося на поверхности Земли и близ нее, лишь несколько долей процента обращается в биосфере. Подавляющая часть углерода сосредоточена в неорганических соединениях (преимущественно в карбонатах) и в каустобиолитах.
Человек участвует в круговороте кислорода за счет дыхания, питания, выделения, использования органического топлива.
39. Границы ареалов и факторы, их определяющие
Границы ареалов могут быть подвижными (транзитивными): прогрессивными (расширяющимися) или регрессивными (сужающимися) и стативными (постоянными). Первый тип границ наблюдается когда вид еще не дошел до естественных границ ареала; второй – когда под влиянием каких-либо факторов площадь ареала сокращается. Стативные границы имеют место в тех случаях, когда вид достиг естественных границ своего ареала.
Положение границ ареалов определяется различными причинами. Границы могут быть эдафическими, если за пределами этих границ вид не находит благоприятных для его существования эдафических условий; конкурентными, когда вид дошел до пределов, на которых он встречается с близким более конкурентоспособным видов, останавливающим его на пути расселения; климатическими, когда за пределами ареала климатические условия для вида становятся неблагопрятными; импедитными, или непроходимыми, когда вид дошел до тех пределов, за которые он не может проникнуть механически.
Границы ареала могут быть пульсирующими, если они ежегодно расширяются или сужаются на некоторое расстояние. Пульсация границ связана с изменениями условий существования на окраине океана.
40. Вертикальная структура биоценоза. Ярусность наземная и подземная
Любое сообщество характеризуется определенной пространственной структурой, связанной в первую очередь с его неподвижными комопнентами – растениями. Вертикальная структура сообщества выражается в его ярусности.
Ярусность может быть надземной и подземной, в сообществах гидрофитов – надводной, подводной и внутригрунтовой. Члены сообщества от верхней части верхнего надземного яруса до нижней части подземного яруса связаны друг с другом. Столб воды в океане над значительными глубинами нельзя считать принадлежащим к одному сообществу, т.к. организмы, обитающие на разных глубинах, характеризуются абсолютно несходными экологическими особенностями.
Ярусы могут быть постоянными, если они построены деревьями или кустарниками с многолетними стволами. Молодые растения при этом не сразу попадают в ярус, образованный взрослыми растениями того же вида. Ярусы, образованные травянистыми растениями, не постоянны. Травы в период вегетации постепенно развиваются, пока не достигнут предельной для себя высоты, при которой они цветут и плодоносят. Индивидуальная изменчивость представителей одного и того же вида приводит к тому, что верхняя граница яруса, образованного травами, представляет собой волнистую линию.
Не все растения оказываются ярусообразующими. Имеются межъярусные виды, входящие в состав того или иного яруса для того, чтобы подняться в наиболее благоприятные для них по условиям освещенности участки. К ним относятся эпифиты (поселяются на других растениях) и лианы (тянутся в кроны растений с различных ярусов). Животные не приурочены постоянно к какому-либо ярусу.
Наземные ярусы в широколиственном лесу (любимый вопрос А. И. Маркина):
- первый древесный ярус – дуб, бук, граб, липа;
- второй древесный ярус – рябина, яблоня, груша;
- подлесок (кустарники) – лещина, бересклет, жимолость;
- травянистый ярус (2 яруса) – осока волосистая, сныть, мятлик дубравный.
41. Первичные сукцессии. Стадии формирования биоценозов
См. вопрос №19.
42. Биогенный круговорот азота
Содержание азота в атмосфере приблизительно равно 70%. Однако для того, чтобы он был использован, он должен быть фиксирован в виде определенных химических соединений.
Фиксация азота происходит в процессе вулканической деятельности, при грозовых разрядах в атмосфере в процессе ее ионизации, при сгорании метеоритов. Однако несравненно большее значение в процессе фиксации азота имеют микроорганизмы: свободноживущие (аэробные бактерии, анаэробный клостридиум Пастера) и обитающие на корнях и листьях растений в особых клубеньках. Процесс фиксации азота требует значительных затрат энергии.
При гниении органических веществ значительная часть содержащегося в них азота превращается в аммиак. Под влиянием живущих в почве нитрифицирующих бактерий аммиак окисляется в азотную кислоту. Азотная кислота взаимодействует с находящимися в почве карбонатами, образуя нитраты. Некоторая же часть азота всегда выделяется при гниении в атмосферу. Кроме того, существуют денитрифицирующие бактерии, отнимающие азот у нитратов, разрушая их с выделением свободного азота или закиси азота N2O.
Таким образом, часть (1) азота сразу попадает обратно в почву в виде нитратов, а часть (2) улетучивается в атмосферу или попадает в гидросферу. Эти потери (2) компенсируются образованием оксидов азота при электрических разрядах, потреблением азота микроскопическими растительными организмами воды (после их отмирания азот попадает в глубокие слои океана) и фиксацией азотобактериями.
Растения же усваивают соединения аммония, нитриты и нитраты из почвы (1), формируя на их основе белки. Эти соединения при поедании растений переходят в тело консументов и в нем преобразуются. Наземные животные, затрачивая энергию, связывают легкорастворимый аммиак в мочевину или в труднорастворимые вещества – гуанин, мочевую кислоту. В печени идет дезаминирование аминокислот, в результате чего и образуется аммиак. Таким образом осуществляется биогенный круговорот азота.
Самым крупным вмешательством человека в круговорот веществ в природе, является промышленная фиксация азота. Для поддержания равновесия в круговороте азота необходимо искусственное развитие процессов денитрификации.
43. Флуктуации биоценозов
См. вопрос №18.
44. Границы ареалов видов и факторы, их определяющие
См. вопрос № 39.
45. Понятие “структура ареала”
В пределах ареала любого вида не все пространство равномерно и сплошь заполнено соответствующими организмами. В одних частях ареала они концентрируются в большем количестве, в других их меньше или же вообще нет. Отсутствие представителей определенного вида на каком-то участке ареала может быть временным и случайным, временным и закономерным. Оно может быть и постоянным, связанным с тем, что пригодные для вида местообитания в данном участке отсутствуют.
При уменьшении численности видов в ареала плотность постепенно уменьшается и в местах с изначально малой плотностью происходят разрывы (дизъюнкции) ареала (более 100 км).
Сплошной ареал – ареал, все участки которого доступные для особей вида, занимающего этот ареал; перенос вида с одного участка на другой осуществляется при помощи современных естественных факторов расселения, причем этот перенос не носит случайного характера и существование вида в разных участках ареала обеспечено. Иными словами, внутри сплошного ареала отсутствуют непреодолимые преграды, разделяющие его на отдельные участки. Дизъюнктивный (разорванный) ареал – ареал, между отдельными частями которого существуют непреодолимые преграды.
46. Моря и океаны как среда жизни
Один из существенных факторов, определяющих жизнь обитателей океана – давление, возрастающее с глубиной. На дне Марианской впадины оно составляет 1100 атм. Известные многочисленные эврибионтности (пофигизм) видов по отношению к давлению (морской еж, некоторые креветки и др.).
Температура вод Мирового океана – один из важнейших факторов распределения организмов. Изменения температур от места к месту в океане более постепенны, чем на суше. Амплитуда температур в тропических широтах - 2°, в умеренных – 6-10°. Поэтому для тропических широт характерно присутствие теплолюбивых стенотермных (кораллы, многие моллюски) форм. В умеренных и приполярных широтах встречаются холодолюбивые стенотермные организмы (белуха, нарвал, сайка, многие тюлени). Многие эвритермные распространены от тропических до приполярных широт. Особенно обильны они в водах умеренного пояса.
Температура воды понижается с глубиной. Глубже 3000 м температура обычно остается постоянной – около 1,8°. Ниже 1500 м вода практически неподвижна. С низкими температурами приполярных вод связано образование на них ледового покрова. У некоторых обитателей этих областей – пагофилов, на ледяных полях проходит вся жизнь.
В некоторых областях наблюдается подъем холодных вод на поверхность – апвеллинг, связанный со сгонами теплых поверхностных вод пассатами. При этом повышается содержание кислорода, увеличивается количество и разнообразие организмов.
Для жизни обитателей океана несомненно велико значение солености его вод. Средняя соленость вод Мирового океана составляет 35 промилле. Среднегодовая соленость колеблется от 30 промилле в приполярных областях до 37,9 промилле в области пассатов. В приполярных областях весной при таянии льдов соленость может опускаться до 15 промилле. Окраинные и внутренние моря могут иметь соленость, которая существенно отличается от солености океана.
Содержание кислорода в воде океанов довольно однородно, хотя может падать с глубиной ниже средней нормы. В некоторых внутренних морях на глубинах падает содержание кислорода и увеличивается количество углекислоты или даже сероводорода (Черное море). Это не позволяет многим организмам обитать здесь.
Движение океанических вод происходит преимущественно в форме течений и затрагивает слой до глубины 1500 м. Течения способствуют перемещению организмов и, бесспорно, играют важную роль в расширении их ареалов. Иногда течения вырывают и перемещают в определенные районы водоросли. Одним из таких районов является Саргассово море. Водоросли при этом еще долгое время живут и служат пищей для специфических организмов. Волновое движение, прибой иногда играет разрушительную роль для некоторых организмов (прибрежных).
Сила света уменьшается с глубиной, ниже 200-500 м свет не проникает. С глубиной увеличивается доля ультрафиолетовой части спектра, поэтому зеленые и бурые водоросли сменяются здесь красными.
47. Развитие ареалов во времени. Дизъюнктивные ареалы
Каждая систематическая категория (вид, род, семейство) характеризуется определенной областью распространения – ареалом. В пределах ареала вид бывает распространен по свойственным ему местообитаниям. Ни один вид не образует сплошного покрова на площади своего ареала.
Сплошной ареал – ареал, все участки которого доступные для особей вида, занимающего этот ареал; перенос вида с одного участка на другой осуществляется при помощи современных естественных факторов расселения, причем этот перенос не носит случайного характера и существование вида в разных участках ареала обеспечено. Иными словами, внутри сплошного ареала отсутствуют непреодолимые преграды, разделяющие его на отдельные участки. Дизъюнктивный (разорванный) ареал – ареал, между отдельными частями которого существуют непреодолимые преграды. Если разорванные участки заселены одной и той же формой вида, то это однородная, или гомогенная, дизъюнкция; если они заселены разными подвидами, разными видами одного рода, разными родами одного семейства, то это разнородная, или гетерогенная дизъюнкция. Гомогенные разрывы ареалов, как правило, моложе гетерогенных. Чем выше гетерогенность, тем более древним является разрыв.
В соответствии с теорией эволюции, вид возник в одном месте и затем расселялся до тех пор, пока не встретил препятствий к расселению. Поэтому любой ареал первично является сплошным. Последующий разрыв может быть обусловлен: 1) вызванным климатическими причинами исчезновением на определенной площади сообществ, с которыми были связаны данные виды растений и животных; 2) истреблением вида на части его ареала человеком или вследствие изменений климата; 3) изменением уровня моря; 4) образованием молодых горных систем. Так, например, плейстоценовые оледенения привели к разрыву ареалов многих широколиственных пород, которые сохранились только в так называемых убежищах, или рефугиумах. Впоследствии наметилась тенденция к смыканию их ареалов.
Для многих морских и наземных обитателей характерны биполярные разрывы ареалов, при которых они присутствуют в бореальных (умеренно холодных) областях Северного полушария и соответствующих им регионах Южного, при этом они отсутствуют в приполярных широтах. Такие разрывы объясняются смыканием ареалов во время похолоданий климата. Для многих океанических видов также характерно амфибореальное (вдоль западного и восточного побережий бореальных частей Тихого и Атлантического океанов). Такие разрывы связаны с теплыми периодами, когда ареалы данных видов охватывали все бореальные области океанов. Есть Американско-Восточноазиатские разрывы некоторых видов, обусловленные образованием Берингова пролива.
При наличии тесных связей между видами исчезновение одного вида в части своего ареала влечет за собой разрыв ареала и другого (хищника, паразита). Эти разрывы, как правило, связаны с изменениями обстановки по сравнению с той эпохой, когда вид возник, развивался и занял определенный ареал.
Первоначальная сплошная область распространения вида (целая популяция) называется центром происхождения вида. Центр происхождения вида часто приурочен к горным областям, где большая разнородность территории и, следовательно, большая изменчивость.
Часть ареала, в которой изменчивость данной систематической группы является наибольшей, называется центром многообразия форм. Как правило эти центры совпадают, т. к. в там, где вид существует дольше, он имеет больше возможностей видоизменяться. Однако при значительном изменении условий существования, центр многообразия форм может существенно отличаться от центра происхождения вида. Согласно миграционной теории возникновения ареала, вид расселяется из центра происхождения до тех пор, пока не достигнет пределов, далее которых он распространиться не может.
Популяции видов крайне неоднородны. Им свойственна генотипическая изменчивость, связанная с потоком генов и их перекомбинацией. При взаимодействии генотипов со средой возникают фенотипы, т. е. группы особей, различающихся по морфологическим, физиологическим и экологическим особенностям. Географическая изменчивость генотипов и фенотипов – результат географической изменчивости условий существования популяции вида. Изменчивость популяций может носить клинальный (постепенный характер). Популяция может представлять и совокупность четко изолированных групп-изолятов. Генетическая изменчивость и свойства популяций обычно различны в центре и на периферии ареала.
У разных систематических категорий организмов тип видообразования разный. Для животных наиболее характерно аллопатрическое видообразование, т. е. возникновение новых видов в географических изолятах. У растений также часто встречается симпатрическое видообразование, т. е. на большей или меньшей площади ареала материнского вида. Заметную роль в видообразовании играет и гибридизация.
48. Высотная поясность в горах. Понятия “высотный пояс”, “тип поясности”
Растительный покров и животное население гор характеризуются сменой поясов от подножья горного хребта до его вершины или до верхних пределов жизни в том случае, когда вершина поднимает за пределы возможности постоянного существования организмов. Высотная поясность – закономерная смена природных ладшафтов снизу вверх в зависимости от изменения факторов среды в связи с высотой над уровнем моря. Идею зональности и поясности, как “общегеографического закона” впервые выдвинул В. В. Докучаев.
Полосы растительного и животного населения в горах называются поясами. Высотный пояс растительности – более или менее широкая и однообразная горизонтальная полоса растительности в горах, составленная или из одного типа растительности или из закономерно чередующихся нескольких. Это же определение можно отнести к биоценотическому, биомному поясу (оробиому). Основные отличия высотных поясов от широтных зон – малая протяженность первых, отсутствие плакорных условий в горах. Тем не менее, подобно интра- и экстразональным биоценозам на равнинах, в горах выделяют внепоясные и межпоясные сообщества.
Горные системы, расположенные в различных зонах, имеют различный набор высотных поясов. Тип поясности – определенный набор поясов растительности (включая основные компоненты биогеоценозов), представленный в определенной последовательной смене. Закономерности в изменении поясности наблюдаются с севера на юг в зависимости от изменения широты, а также связаны с изменениями степени сухости и континентальности климата. Иногда выделяют ряды поясности, сходные по условиям увлажнения, но различающиеся по температурному режиму.
50. Эндемики. Палео- и неоэндемичные ареалы
Под эндемичными видами (эндемами) понимаются виды, ареалы которых не выходят за пределы какой-либо естественной области, ограниченной от соседних. Обычно верхним пределом размеров такой области является биотическая провинция.
Видовой эндемизм особенно характерен для горных вершин, резко отграниченных от равнинных пространств или других горных вершин; островов, ранее составлявших части континента, ныне изолированных водой от остальной суши; участков, отграниченных природными условиями от других сходных участков (например, островные березовые лески полупустынь Казахстана, в которых произрастает киргизская береза).
Эндемы могут иметь различный возраст. Древние эндемы, или палеоэндемы, связаны с филогенетической обособленностью и эколого-географической изолированностью (приурочены горным системам, древним островам). Палеоэндемами также являются виды, сузившие свой ареал в результате изменения обстановки. Молодые эндемы, или неоэндемы, обычно появляются там, где какая-либо более древняя форма в изменившихся условиях обитания дает начало более молодой. Часто это не виды, а подвиды и популяции. Палеоэндемы не имеют близких таксономически видов в других областях, неоэндемы имеют.
53. Охрана редких видов растений и животных. “Красные книги”
Человек существенным образом влияет на сообщества живых организмов непосредственно уничтожая их или изменяя условия их существования. Для того чтобы сохранить природу нашей планеты, необходимо, чтобы каждый проникся убежденностью беречь обитателей Земли!
При использовании природных ресурсов, в том числе возобновимых ресурсов биосферы, необходимо сохранение экологического разнообразия как в пределах культурного ландшафта, так и путем оставления эталонов совершенно нетронутой дикой природы (принцип поляризованной биосферы).
Заповедники, национальные парки, заказники (ограниченное использование природных ресурсов) и другие резерваты дикой природы часто выполняют функции центров стабильности данного участка биосферы, благотворно воздействуют на природу далеко за пределами своих границ. Они служат хранителями естественных сообществ генетического фонда.
В связи с настоятельной необходимостью сохранения и слежения за естественными экосистемами возникла идея о биосферных заповедниках. Отличительными их чертами должны быть: 1) обеспечение естественной регуляции типичных экосистем (это требует большой площади; 2) представленный набор экосистем должен обладать эталонной ценностью; 3) научные исследования должны быть обеспечены квалифицированными исполнительными кадрами.
С целью охраны редких видов в 1948 г. был создан Международный союз по охране природы (МСОП). При МСОП существует также Фонд дикой природы (ВВФ). При участии этих организаций была создана Международная красная книга МКК (Red Data Book). Виды, занесенные в МКК, разделяются по категориям опасности:
- на грани исчезновения
- сокращающиеся
- редкие
- неопределенные
- востановленные
Для всех видов указываются данные об ареале, а также рекомендуемые меры по охране. В качестве приложения к МКК существует черный список исчезнувших после 1600 г. видов.
54. Эндемики и эндемичные ареалы
См. вопрос №50.
57. Зональные, интра- и экстразональные биоценозы. Их место в структуре растительного покрова суши
В пределах каждой природной зоны размещение экосистем контролируется почвенными и геоморфологическими условиями. В связи с этим используются такие понятия, как зональные, интразональные и экстразональные биоценозы.
Зональный биоценоз занимает типичные для данной зоны так называемые плакорные участки (плоские или слабоволнистые, слабонаклоненные, суглинистые, хорошо дренируемые поверхности в пределах междуречий).
Интразональные и экстразональные биоценозы приурочены к специфическим местообитаниям, где экологические условия отличаются от местообитаний плакоров. Наличие таких ландшафтов обычно связано с повышенным грунтовым увлажением или с особыми эдафическими условиями. Интразональные ландшафты также зональны, но их зональность своеобразна (например, зональность болот в Западной Сибири).
Интразональные биоценозы нигде не образуют своей зоны, но включены в одну или несколько природных зон во внеплакорных условиях (например, пойменные луга, биоценозы болот).
Экстразональные биоценозы – участки зональных биоценозов виде островков, включенных в другие природные зоны во внеплакорных условиях (обычно на склонах определенной экспозиции).
58. Климакс. Представление об устойчивости биоценозов
См. вопрос №12.
59. Роль человека в формировании ареала
Действия человека могут вызвать сужение границ ареала путем прямого уничтожения вида на значительной части ареала (в основном – промысловые животные). Примеры: бурый медведь, бизон, россомаха, глухарь. Некоторые виды полностью исчезли: дрофа, степной сурок.
Человек может создавать преграды для расселения видов. Искусственное преодоление преград с помощью человека также возможно. Например, кролики и опунции (кустарник семейства кактусовых) были привезены человеком в Австралию, олени – в Новую Зеландию.
Изменения условий среды из-за деятельности человека также могут вызвать расселение видов, расширение первоначального ареала. Поэтому многие сорные и мусорные растения (обыкновенный одуванчик, пастушья сумка), а также животные-синантропы (серая и черная крысы, домовая мышь, домовый воробей) имеют космополитные ареалы.
60. Основные различия между биоценозами песчаных и глинистых пустынь
1. Глинистые пустыни – основной зональный тип пустынной растительности. Они характерны для северных пустынь. Песчаные пустыни распространены, как правило, в более южных районах.
2. Для глинистых пустынь характерна флористическая бедность, песчаные пустыни в этом отношении наиболее богаты из всех пустынь.
3. Глинистые пустыни характеризуются большой задернованностью, т. е. большим проективным покрытием
4. В глинистых пустынях верхний ярус обычно представлен деревьями (черный саксаул). В песчаных пустынях деревьев почти нет, встречаются деревца (песчаная акация, эремоспартон) и кустарники (джузгун, белый саксаул).
5. В растительном покрове глинистых пустынь преобладают полукустарнички (полынь, биюргун) и многолетние травы. На севере встречаются дерновинные злаки, эфемеров мало. Песчаные пустыни характеризуются преобладанием травянистых растений – эфемеров (костры, маки, злак селин) и эфемероидов (осока песчаная).
6. Глинистые пустыни – основной зональный тип пустынной растительности. Они характерны для северных пустынь. Песчаные пустыни распространены, как правило, в более южных районах.
7. Для глинистых пустынь характерна флористическая бедность, песчаные пустыни в этом отношении наиболее богаты из всех пустынь.
8. Глинистые пустыни характеризуются большой задернованностью, т. е. большим проективным покрытием
9. В глинистых пустынях верхний ярус обычно представлен деревьями (черный саксаул). В песчаных пустынях деревьев почти нет, встречаются деревца (песчаная акация, эремоспартон) и кустарники (джузгун, белый саксаул).
10. В растительном покрове глинистых пустынь преобладают полукустарнички (полынь, биюргун) и многолетние травы. На севере встречаются дерновинные злаки, эфемеров мало. Песчаные пустыни характеризуются преобладанием травянистых растений – эфемеров (костры, маки, злак селин) и эфемероидов (осока песчаная).
61. Пелагиаль как экологическая область океана
См. вопрос №9.
62. Голарктическое флористическое царство. Восточноазиатская область
Голарктическое флористическое царство охватывает большую часть Евразии (в том числе всю территорию России), Северную Африку и Северную Америку – больше половины всей суши. Для него характерно обилие эндемичных родов и эндемичных семейств (30) с очень малыми ареалами. Многие ареалы разорваны из-за деятельности плейстоценовых ледников. Голарктическое царство подразделяют на подцарства: Бореальное (области Циркумбореальная, Восточноазиатская, Атлантическо-Североамериканская и область Скалистых гор), Древнесредиземноморское (области Макаронезийская, Средиземноморская, Сахаро-Аравийская, Ирано-Туранская), Мадреанское (Сонорское). Последнее охватывает большую часть Мексики и юго-запад США.
Восточноазиатская область включает большую часть Гималаев, части северо-восточных пограничных районов Индии, значительную часть континентального Китая, Приморье, значительную часть бассейна Амура, юго-восточное Забайкалье, восточную Монголию, Корею, Тайвань, Южные Курилы, южный Сахалин.
Флора восточноазиатской области весьма богата: включает 14 эндемичных семейств и более 300 эндемичных родов. Эндемичны из хвойных семейства гинкговых, цефалотаксовых; многие покрытосеменные. О древности флры свидетельствует, помимо обилия энедмиков, наличие весьма примитивных растений – магнолиевых, лютиковых и гамамелидовых. Эта область представляет собой один из главных центров развития высших растений Голакртики и Палеотрописа и гигантское убежище древних форм.
63. Важнейшие изменения растительности и животного населения тайги с севера на юг
В направлении с севера на юг в тайге выделяют широтные подзоны: северная, средняя и южная тайга. Для северной тайги характерны низкорослые древостои с небольшой сомкнутостью крон, представляющие собой переход к тундровым редколесьям. Под изреженным древесным пологом обычно развит ярус из низкорослых субарктических кустарников (берез, ив). Напочвенный покров образован мхами и лишайниками. На равнинах наблюдается сильная заболоченность.
По мере движения к югу древостой становится более высоким, увеличивается роль травяно-кустарничковой растительности. Леса приобретают более сложную структуру, возрастает сомкнутость крон. Площадь болот и заболоченных лесов при этом сокращается. Хорошо развиты травяно-кустарничковый ярус и моховой напочвенный покров (лишайников мало).
Южнотаежные леса высокоствольны, велика сомкнутость крон, флористический состав богаче, напочвенный покров слабый. В европейской южной тайге появляются широколиственные породы. В состав подлеска и травяного покрова входят виды, характерные для широколиственных лесов.
Наиболее хорошо широтная зональность выражена в тайге Западной Сибири.
64. Характеристика листопадных тропических лесов
В области распространения листопадных тропических лесов количество осадков равно 800-1300 мм, продолжительность сухого периода составляет 4-6 месяцев.
В этих лесах, несмотря на название “листопадных”, произрастают главным образом в нижних ярусах значительное количество вечнозеленых деревьев. Однако их здесь меньше, чем в полулистопадных. Обычны деревья со сложными листьями. Деревья, как правило, корявы и невысоки. Основную массу составляют деревья нижнего яруса, не превышающие 12 м. Имеются и поднимающиеся над общим уровнем древостоя деревья-эмердженты до 20, иногда до 40 м высоты.
Кустарниковый ярус сомкнут; травяной покров почти отсутствует. В более светлых участках леса в травяном покрове обильны злаки. Среди эпифитов отмечены орхидеи и папоротники. По деревьям и одна по другой вьются лианы.
Тиковые леса характеризуются тем, что тик, образующий верхний древесный ярус, сбрасывает листву, но среди деревьев нижнего яруса имеются и виды вечнозеленые. Саловые леса образованы сбрасывающим листву салом. В подлеске также встречаются деревья, сохраняющие на сухой период листву.
65. Важнейшие особенности тундровых биоценозов. Жизненные формы, структура, изменчивость.
Зона тундры имеет циркумполярное распространение и занимает 14% площади СССР. Осадков в тундре выпадает мало: 200-300 мм в год, но из-за низких температур коэффициент увлажнения близок к 1. Большая часть осадков приходится на лето, поэтому снежный покров маломощен. Лето короткое, зима очень длинная и суровая. Из-за низкой температуры воздуха высока относительная влажность. Сильный ветер сдувает снег в понижения. Почвы мерзлые, летом образуется деятельный слой. Происходят солифлюкиця, децерация, активное заболачивание (60% территории тундры – болота). Из-за низких температур почвообразовательные процессы развиты слабо. При недостатке кислорода идет активное оглеение. Почвы бедны элементами минерального питания.
Вегетационный период очень короткий. Южная граница тундры совпадает с изотермой июля +10°С, поэтому деревья здесь расти не могут. У немногочисленных цветковых растений все-таки успевают созревать плоды (благодаря наличию полярного дня).
Тундра флористически и фаунистически бедный биом. Растений всего 250-300 видов, некоторые эндемичны. Фитомасса не превышает 300 ц/га. Растительность тундры характеризуется отсутствием деревьев и преобладнием во многих типах тундры мхов и лишайников. Однолетников мало, т. к. мала продолжительность вегетационного периода и малы его температуры. Из многолетников много вечнозеленых форм. Много также кустарников с невысокими, стелющимися по поверхности почвы деревянистыми стволиками и ветвями, прижатыми к поверхности, а также травянистых дерновинных растений. Все эти формы экономят тепло, прижимаясь к земле.
Из зимнезеленых кустарничков наиболее широко распространены куропаточья трава, кассиопея, брусника, вороника; из кустарничков с опадающей листвой – черника, карликовая березка, карликовая ива. Промерзание почвы препятствует развитию растений с подземными запасающими органами.
Оседлых птиц в тундре практически нет, из пресмыкающихся встречается только одна живородящая ящерица, амфибий – 3 вида. Многолетняя мерзлота и сильное заболачивание ослабляют роющую деятельность животных. Среди млекопитающих наиболее типичными являются лемминги, из птиц характерные водные и околоводные. Эндемичных видов животных очень мало.
Зимой большинство пойкилотермных животных впадает в анабиоз, а гетеротермных – в спячку (длиннохвостный суслик, черношапачный сурок). Гомеотермные животные обычно зимой покидают зону тундры, откочевывая на юг (белая сова, пуночка, северный олень) или улетая не теплые зимовки. Млекопитающие и птицы, активные зимой в тундре, имеют густой шерстный покров, накапливают толстый слой подкожного жира. Открытость ландшафта обусловливает стайный и колониальный образ жизни водоплавающих птиц и стадность оленей.
Среди трофических групп животные доминируют сапрофаги. Значительные колебания численности одних животных приводят к крупным флуктуациям всех биоценозов.
66. Изменение состава и структуры степных биоценозов с севера на юг
На границе с широколиственными лесами располагаются луговые степи, которые обычно относят к лесостепной зоне, где на водоразделах сочетаются леса и красочные степи.
В северных степях развиты разнотравно-дерновинно-злаковые или красочно-ковыльные степи. В фитоценозах северных степей большую роль играют ковыли, преимущественно перистые, создающие серебряный аспект при цветении. Среди других дерновинных злаков распространены типчак и тонконог. В северных степях наблюдается многократная смена аспектов.
Подзона южных степей характеризуется дерновинно-злаковыми или бескрасочно-ковыльными сообществами. Во флоре увеличивается количество типичных ксерофитов, склерофильных злаков, разнотравья. Красочное разнотравье сменяется бескрасочным сухим. Увеличивается роль эфемеров и эфемероидов. Аспектов меньше, чем в северных степях, раньше наступает период покоя, травостой изреженный.
В самой южной части степной зоны развиты опустыненные степи, в которых кроме дерновинных злаков (типчак, ковылки, змеевка) присутствуют различные виды полыни, а также кустарники – караганы, спиреи. На засоленных местах встречаются пустынные растения – биюргуны, солянки, мощный злак чий. Растительность очень разрежена.
67. Географические особенности размещения темнохвойных и светлохвойных лесов России.
В Европе светлохвойные породы приурочены преимущественно к почвам легкого механического состава или сменяют темнохвойную тайгу после пожаров, вырубок. Такие леса образованы преимущественно обыкновенной сосной.
В Сибири и Северной Америке первичные светлохвойные леса могут быть связаны с почвами более тяжелого механического состава. Здесь играют большую роль различные виды лиственниц. Лиственница является светолюбивой породой и выносит очень низкие температуры и большие амплитуды. В Западной Сибири распространена лиственница сибирская, причем она перемежается с небольшими массивами темнохвойных и сосновых лесов, в Восточной – менее прихотливая, преимущественно горная порода, - лиственница даурская.
На северо-востоке Европейской части, на Урале и в Сибири встречаются кедровые леса. Кедр достаточно неприхотлив, но предпочитает хорошо дренируемые почвы.
68. Неотропическое флористическое царство. Амазонская и Андийская области
Неотропическое царство занимает тропическую Южную и Центральную Америку, а также южную оконечность Флориды. Имеется 25 эндемичных семейств (настурциевые и др.). Большое сходство с Палеотропическим царством (семейства лавровых, крапивных, миртовых, мальпигиевых, бигнониевых, арековых) свидетельствует о том, что флора Нового Света уходит корнями в старый Свет. Общих родов более 450. Ряд семейств (пассифлоровые, миртовые и др.) распространен в Неотрописе шире, чем в Палеотрописе. Ряд почти эндемичных семейств Неотропического царства имеет по одному или по несколько представителей в Западной Африке, что свидетельствует о связи между этими царствами до сравнительно недавнего времени (начала третичного периода). Таковы семейства бромелиевых, кактусовых, рапатовых. Голарктическое царство включает Карибскую, Гвианского нагорья, Амазонскую, Бразильскую и Андийскую области.
Амазонская область – очень богатая флористически область. Здесь произрастают влажные тропические леса. Семейственный эндемизм не велик, однако много эндемичных родов, например, гевея.
Андийская область – самая высокогорная в царстве. Почти до экватора доходит распространение некоторых бореальных Голарктических и Голантарктических видов. Здесь произрастает известное своими целебными свойствами хинное дерево, много эндемичных видов картофеля и помидора.
69. Характеристика биома саванн
Саванны – тип растительности тропического пояса, обычно древесно-кустарниковый, но иногда почти лишенный древесного яруса. Дождливый сезон здесь обычно один, сменяется сухим продолжительностью 4-6 месяцев. Эти смены влажного периода сухим создают своеобразие условий существования животных и растений.
Деревья часто имеют толстую кору с мощным слоем пробки. Они сбрасывают на сухое время года листья. Многие деревья имеют зонтиковидную крону, способствующую равномерному распределению влаги на площади, занятой поверхностными корнями, а также уменьшающую иссушающее действие ветра в сухой период. Травяной покров различен – во влажных условиях он образован высокими злаками, в сухих саваннах – низкими злаками и различными полукустарниками, вместе со злаками образующими сомкнутый травяной покров. Деревья или равномерно распределены среди травостоя наподобие фруктового сада или образуют рощицы, чередующиеся с участками, занятыми травами. С началом засухи надземные части трав высыхают, с деревьев опадает листва. В конце периода засухи обычно деревья саванн цветут, а с началом влажного периода одеваются листьями.
Для всех саванн характерно обилие стадных мелкопитающих: антилопы, зебры, слоны, жирафы а Африке; мелкие олени в Южной Америке; сумчатые, в том числе гигантский кенгуру в Австралии. Из бескилевых птиц в Африки водится африканский страус, в Австралии - эму, в Южной Амерке – нанду.
Во всех саваннах, кроме австралийских, много землероев-грызунов. В Африке также преобладают трубкозубы. В Австралии роющие плацентарные млекопитающие заменены сумчатыми – вомбатом, сумчатым кротом и др. Термиты устраивают в саваннах плотные глинобитные постройки, которые могут разрушаться со съедением всех обитателей африканским трубкозубом.
Обилие крупных копытных и других травоядных животных – причина существования в саваннах значительного количества хищников: львы и гепарды в Африке, ягуары в Южной Америке, собака динго в Австралии. Также характерны млекопитающие и птицы падалееды – гиены, грифы, стервятники, кондоры. У крупных птиц-падалеедов существует система оповещения о наличии корма.
Источниками водопоя для обитателей саванн являются реки, которые протекают по долинам, поросшим так называемыми галерейными лесами. Здесь в условиях значительной влажности воздуха обитают различные кровососущие двукрылые насекомые, например, муха-цеце. Насекомые переносят опасные для животных или человека болезни: нагану, сонную болезнь, болезнь Чагаса.
70. Растительность и животное население степей
Зона степей представлена в Евразии степями, в Северной Америке прериями, в Южной Америке – пампасами, в Новой Зеландии – сообществами туссоков. Для степей характерен хороший обзор, обилие растительной пищи, относительно сухой летний период, существование летнего периода покоя, или полупокоя. Биомасса степей 1500-2500 ц/га.
Среди преобладающих жизненных форм растений степи выделяются дерновинные злаки (туссоки) или корневищные злаки, не образующие дерновин. Распространена особая форма перекати-поле (переносятся ветром и разбрасывают семена). Среди двудольных травянистых растений выделяются две группы – северное красочное разнотравье, представленное мезофильными растениями с крупными яркими цветами или соцветьями, и южное бескрасочное разнотравье, состоящее главным образом из неярких, малозаметных ксерофитов.
Характерны для степей однолетние эфемеры, отцветающие весной, после цветения отмирающие, и многолетние эфемероиды, у которых после отмирания надземных частей остаются клубни, луковицы, подземные корневища. Безвременник развивает листву весной, а осенью цветет. На поверхности почвы растут ксерофильные мхи, кустистые и накипные лишайники, иногда сине-зеленые водоросли. Для степи также характерны растущие группами или одиночные кустарники.
В степи наблюдается многократная смена аспектов, т. е. изменение внешнего облика степей в связи с тем, что цветущие растения, обычно развивающиеся в массах, сменяют друг друга. Реже аспекты создаются массовыми видами животных, они обычно меняются в течение дня.
В степи широко распространен норный образ жизни. Норы служат для укрытия, спячки, в качестве кормовых ходов (некоторые животные питаются подземными частями растений). Роющая деятельность степных животных играет большую роль в изменении характера почвенного покрова и т. п. Многие строящие норы животные ведут колониальный образ жизни (например, суслики). Для колониальных животных существенны звуковые предупредительные сигналы. В норах могут жить переносчики различных болезней – эктопаразиты. Животные, не устраивающие нор, обычно ведут стадный образ жизни (сайгак и др.).
Ряд видов впадает на лето в спячку, которая потом переходит в зимнюю. У мелких грызунов (полевки) и у некоторых насекомых имеют место массовые размножения. В эти периоды уничтожаются основные виды кормовых растений, животные вынуждены мигрировать.
71. Важнейшие особенности биоценозов темнохвойных таежных лесов
Продолжительность периода со среднимесячными температурами выше 10°С составляет 2 – 4 месяца. Число ярусов в темнохвойных лесах обычно равно двум-трем. Кроме древесного яруса, могут быть развиты травяной или травяно-кустарничковый ярусы и моховой в тех случаях, когда лес не является мертвопокровным. Иногда отсутствует и травяной ярус. Кустарники единичны и выраженного яруса не образуют.
Лесная подстилка разлагается медленно, поэтому некоторые травянистые растения не образуют хлорофилла и питаются сапрофитно. Освещение в течение всего года одинаковое, поэтому нет растений, приурочивающих развитие цветков к ранневесенним месяцам. Венчики цветков белые или бледно окрашенные. Травы и кустарнички размножаются обычно вегетативным путем, образуя куртины, группы (из-за сильного затенения). Токи воздуха очень слабые, поэтому семена многих растений очень легкие (грушанки, орхидеи). Для развития зародышей таких растений необходимо участие гриба – микориза (мутуализм).
Перенос многих семян осуществляется животными, поедающими сочную мякоть плодов (черника, брусника, толокнянка), высокая кислотность сока ягод препятствует развитию семян в нетронутой ягоде. Также распространение семян может происходить при растаскивании семян муравьями.
В тайге мало стадных животных, поскольку наличие древостоя затрудняет зрительное оповещение об опасности. Иногда встречаются кабаны, заходят волки и северные олени. Основные способы охоты – подкарауливание и скрадывание. Среди хищных птиц особенно характерны ястребы. Сравнительно небольшое количество животных покидают тайгу зимой. Многие способны питаться веточными кормами (лось, заяц). Ряд видов обитает на деревьях, кормится на земле (лесной конек, дрозды), другие – наоборот (тетерев, рябчик, глухарь, тетерев). Некоторые животные питаются семенными кормами (белки, бурундуки, мышевидные грызуны). В годы неурожая они мигрируют обычно на запад, многие гибнут при переправах. Из насекомых, поедающих хвою, широко распространен непарный шелкопряд; вредители древесины – жуки-усачи и их личинки и др. Многие млекопитающие и птицы, пища которых связана с деревьями, хорошо приспособлены к лазанию и нередко живут на деревьях.
72. Важнейшие особенности организации пустынных биоценозов
Пустынный тип растительности объединяет сообщества с господством ксерофитных полукустарничков. В целом, сообществам свойственна низкорослость, флористическая бедность, невысокое проективное покрытие, разреженное произрастание растений, сочетающееся со значительной сомкнутостью подземных частей.
Основной фактор, ограничивающий развитие растений в пустыне – дефицит влаги в почве и воздухе, с этим связано большинство приспособлений. Большинство пустынных растений является ксерофитами. Среди них преобладают полукустарнички (полыни, солянки), кустарники (джузгун, белый саксаул), встречаются деревья (черный саксаул), деревца (песчаная акация), многолетние и однолетние растения, мхи, лишайники. Для ксерофитов характерен незначительный годовой прирост, мелколиственность или безлистность, мощная корневая система.
В пустынных биоценозах также встречаются мезофиты, вегетационный период которых совпадает с коротким периодом выпадения атмосферных осадков (лето в пустынях умеренного пояса, зима или ранняя весна в тропиках). К ним относятся однолетние эфемеры (костры, мортуки) и многолетние эфемероиды (мятлик луковичный, осоки, луковичные), а также водоросли.
Большое влияние на пустынные биоценозы играет субстрат. Избыток солей в почве для некоторых растений является токсичным. Выделяют гликофиты – растения незасоленных почв, галофиты – растения засоленных почв, гликогалофиты и гипсофиты. По механическому составу грунтов пустыни подразделяют на глинистые, песчаные, солончаковые и каменистые; иногда выделяют отдельно лессовые пустыни.
Характерная черта пустынных животных – норный образ жизни. Некоторые быстро передвигающиеся формы (антилопы, птицы рябки) преодолевают неблагоприятно сложившиеся условия добывания корма за счет способности к быстрому передвижению и живут крупными стадами или стаями.
73. Неотропическое флористическое царство. Амазонская и Андийская области
См. вопрос №68.
74. Характеристика степного биома
См. вопрос №70.
75. Характеристика биоценозов тропических влажных лесов
Влажные тропические леса (ВТЛ) произрастают в оптимальных условиях влажности и температурного режима. Эти условия обеспечивают максимальную продукцию растительного покрова, а следовательно, и общую продукцию. Биомасса составляет 3500-7000 ц/га, иногда достигает 17000 ц/га. Климат характеризуется малыми годовыми (1-2°) и несколько большими суточными (9°) амплитудами температур, большим (1000-5000 мм) количеством осадков, высокой (40-100%) относительной влажностью. Продолжительность дня колеблется мало, осадки выпадают преимущественно во второй половине дня и связаны с конвекцией. Часты ураганы, образующие окна в древостое.
В ВТЛ выделяются две группы деревьев – тенелюбивые дриады и светолюбивые номады. При образовании окон в древостое развиваются сначала номады, потом под их пологом вырастают дриады. Поэтому для биоценозов ВТЛ характерна мозаичность.
Почвы красные, красно-желтые и желтые ферралитные, они недостаточно обеспечены микроэлементами, подстилка из листьев небольшая, часто отсутствует. Почвы, как ни странно, бедны водно-растворимыми минеральными соединениями, которые по преимущественно заключены в деревьях.
Для ВТЛ характерно значительное число видов деревьев – от 40 на островах до 170 на материках. Значительно меньше число трав. Из межъярусных растений в ВТЛ много лиан (карабкающиеся с помощью ускиков, цепляющиеся, обвивающиеся, опирающиеся), эпифитов (розетки, гнездовые эпифиты, полуэпифиты, протоэпифиты), имеются деревья-душители (в основном фикусы). Также встречаются небольшие по размерам эпифиты, живущие на листьях – эпифиллы. Они относятся к водорослям, мхам и лишайникам. Для деревьев ВТЛ характерно являение каулифлории или рамифлории – развитие цветков на стволах ниже кроны или на толстых ветвях, где их легче отыскать опылителям.
Смена листьев у разных деревьев происходит по-разному. Иногда отдельные эмердженты могут стоять без листьев несколько дней. Смена листьев может идти непрерывно в течение года, может происходить по-разному на различных побегах, могут чередоваться периоды листообразования и покоя. Деревья могут цвести и плодоносить непрерывно в течение года или по несколько раз в году, некоторые цветут ежегодно или раз в несколько лет.
Вертикальная структура ВТЛ характеризуется единичными деревьями-эмерджентами и плохо выраженной ярусностью деревьев основного полога, особенно при полидоминантной структуре. Причины плохой выраженности ярусов: древность сообщества (высоко совершенное “прилаживание” деревьев разных видов друг к другу) и оптимальность условий существования (велико число видов в биоценозе).
Кустарников почти нет. Мелкие деревянистые растения имеют один ствол и относятся, таким образом, к деревьям: молодым или карликовым. Наличие только одного ствола связано с плохой освещенностью. Для травянистого покрова ВТЛ характерно преобладание одного вида (обычно папоротник или селагинелла) с незначительной примесью других. Встречаются многолетние очень высокие травянистые растения.
Среди обитателей ВТЛ многие связаны с кронами. Это обезьяны, полуобезьяны, ленивцы, белки, летяги, шерстокрылы; из насекомоядных – тупайи, мыши и крысы. Многие млекопитающие имеют приспособления к планирующему полету, много зверей и птиц-дуплогнездников (белки, бурундуки, птицы-носороги и др.). Многие животные строят убежища на поверхности почвы из гниющих листьев (муравьи, сорные куры); термиты живут в подземных гнездах.
Обилие разнообразных видов и жизненных форм приводит к развитию сложных симбиотических отношений. Например, ряд растений имеет пустоты в стволах, где поселяются хищные муравьи, охраняющие растения от муравьев-листорезов. Очень развиты явления покровительственной окраски и формы (например зеленая окраска ленивца, возникающая благодаря живущим в его шерсти водорослям). Также встречается апосематическая окраска (яркая, отпугивающая, указывает на то, что вид несъедобен). Иногда встречаются виды с псевдоапосематической окраской.
76. Краткая характеристика Австралийской фаунистической области
Австралийская область – единственная в составе фаунистического царства Нотогея. Характерными особенностями Австралийской фаунистической области является обилие эндемиков, в том числе однопроходных, наличие некоторых групп сумчатых и почти полное отсутствием плацентарных за исключением грызунов.
Как уже было сказано, характерными являются подкласс однопроходных млекопитающих (утконосовые и ехидновые) и шесть семейств сумчатых, из которых фалангер распространен до о. Сулавеси на севере. Плацентарные, кроме завезенной и одичавшей собаки динго, богато представлены разнообразными грызунами, включающие древние примитивные и специфичные формы. Здесь встречаются виды родов цепкохвостых мышей, кенгуровых мышей, крыс.
Граница Австралийской области с Индо-Малайской проходит по так называемой “линии Уоллеса” между о-вами Бали и Лембок. Остров Сулавеси обычно относят к Австралийской области, иногда его считают особой переходной зоной. В пределах Австралийской области выделяются Папуасская, Австралийская, Новозеландская, Полинезийская и Гавайская подобласти.
77. Растительность и животное население листопадных тропических лесов
См. вопрос №64.
78. Капское флористическое царство
Капское царство занимает южную оконечность Африки. Это самое маленькое из флористических царств, однако его выделение очевидно благодаря крайнему своеобразию флоры. Флора имеет около 7000 видов, включает 7 эндемичных семейств. Из Капского царства произошли более 280 родов, из них более 210 эндемичны для него.
Для Капского царства характерна флора двойственного происхождения. Несомненно наличие значительных связей с южными материками, с которыми юг Африки образовывал Гондвану. Наследниками Гондваны явились представители семейств протейных и рестионовых, например, серебряное дерево (большая часть распространена в Австралии). Ряд других растений гондванского происхождения проник сюда после расчленения Гондваны.
Наряду с видами южного происхождения в Капском царстве много видов, связывающих его с растениями тропической Африки и со Средиземноморьем. В Капском царстве насчитывается 20 эндемичных родов вересковых, множество рутовых. Очень много кустарников, разнообразие стеблевых суккулентов. Граница с Палеотрописом проходит по реке Оранжевая.
Капская флора – неисчерпаемый источник декоративных растений, в том числе луковичных и клубненосных однодольных – амариллисов, кливий, гладиолусов и др.
79. Растительность и животное население саванн
См. вопрос №69.
80. Растительность и животное население биома широколиственных лесов
Широколиственные леса произрастают в условиях более мягкого климата, чем хвойные леса. Биомасса 4000-5000 ц/га (близка к южным хвойным лесам). Широколиственные деревья, в отличие от большинства хвойных, сбрасывают листву на зиму. Поэтому ранней весной под их пологом светло, многие деревья (бук, дуб) цветут одновременно с распусканием листьев; кустарники (лещина, волчье лыко) – до распускания листьев.
Мощная и рыхлая подстилка предохраняет почву от резкого понижения температуры, зимнее промерзание незначительно. В связи с этим многие виды травянистых растений начинает развиваться в конце зимы. Это группа весенних эфемероидов, которые, закончив цветение ранней весной, затем или вегетируют, или теряют свои надземные органы (ветренница дубравная, гусиный лук). Бутоны этих растений зачастую развиваются с осени. Мощная подстилка позволяет перезимовывать и разнообразным беспозвоночным. Поэтому богаче и почвенная фауна (кроты и др.).
Ярусная структура широколиственных лесов много сложнее структуры хвойных лесов умеренного пояса. Обычно выделяются от одного (бучины) до трех (дубравы) ярусов древостоя, два яруса кустарников и два-три яруса трав.
Питательные плоды деревьев, а также раскидистые мощные ветви и большая дуплистость способствуют распространению многочисленных млекопитающих и птиц. У многих видов животных наблюдается специализация в питании (например, дубонос потребляет только семена косточковых древесных и кустарниковых пород). Активны роющие эивотные, например, муравьи, способствующие развитию дернового процесса. В связи с ослаблением ветра в лесах много насекомых с порхающим полетом. Много вредителей леса, в том числе листогрызущих. Некоторые виды деревьев вынуждены даже обновлять листву летом.
Широколиственные леса не образуют сплошной полосы. В Европе с запада на восток каштановые леса сменяются буковыми, а потом дубравами. Отдельные острова липовых лесов встречаются за Уралом – в предгорьях Кузнецкого Алатау и др. На Дальнем Востоке произрастают дуб, клен, маакия, элеутерококк, аралия; в состав подлеска входят жимолость, сирень, рододендрон, бирючина и др. В более южных районах Дальнего Востока обильны лианы (актинидия и др.) и эпифиты. В северо-восточной части Северной Америки имеются леса с доминированием бука американского и сахарного клена, иногда в лесах встречаются лианы – “дикий виноград”. Более континентальные районы приатлантических штатов заняты дубравами.
81. Характеристика Эфиопской области
Эфиопская фаунистическая область занимает большую часть Африки, кроме ее северной части (Сахары), а также Мадагаскар с тяготеющими к нему островами. Фауна включает 37 семейств млекопитающих, из которых 9 – куницы, землеройки, кошки, белки, свиньи, полорогие и др. распространены очень широко. Шесть семейств млекопитающих (человекообразные обезьяны, лемуры лори, слоны, носороги, оленьки, ящеры) объединяют эфиопскую фауну с восточной при отсутствии в Палеарктике. Эндемичны или незначительно выходят за пределы области лемуры, руконожка, прыгунчики, выдровые землеройки, тенреки, златокроты, гиены, шипохвостые белки, даманы, бегемоты, жирафы, трубкозубы.
Для области характерно разнообразие копытных. Велико разнообразие антилоп, обильны зебры, жирафы, носороги, слоны, бегемоты. Полностью отсутствуют семейства кротов, медведей, бобров, тушканчиков, верблюдов, тапиров, оленей. Среди птиц 15 эндемичных или почтиэндемичных семейств: африканские страусы, цесарки, секретари и др. Ряд птиц общи с Индо-Малайской (Восточной) областью: птицы-носороги, павлины. Очень характерны рябчики, дрофы, грифы. Из пресмыкающихся многочисленные вараны, агамы, сцинки. Хамелеоны почти эндемичны. Весьма разнообразны гадюки, питоны, крокодилы. Из амфибий жабы и настоящие лягушки представлены очень широко.
Область разделяется на четыре подобласти – Восточноафриканскую, Южноафриканскую, Западно-Африканскую и Мадагаскарскую.
82. Сравнительная характеристика биоценозов тайги и хвойно-широколиственных лесов
1. Хвойно-широколиственные (смешанные) леса распространены в Евразии только в Европе и на Дальнем Востоке.
2. В смешанных лесах широко распространены широколиственные породы – дуб, липа, клен, граб.
3. Важнейшими лесообразующими породами смешанных лесов являются широколиственные породы – дуб черешчатый и др.
4. В смешанных лесах сочетаются представители бореальной и неморальной (дубравной) флоры во всех ярусах, поэтому больше флористическое разнообразие.
5. Широколиственные породы встречаются в верхнем древесном, 2-м древесном ярусах и в подлеске.
6. Хорошо развит ярус подлеска – лещина, жимолость, бересклет, калина.
7. Травяной покров состоит из таежных видов (кислица, майник, грушанки, папоротники) и растений широколиственных лесов (сныть, осока-волосистая, зеленчук, звездчатка жестколистная, ясменник душистый).
8. Развиты эфемероиды – хохлатка, гусиный лук, ветреница (подснежное развитие).
9. Моховой покров развит слабо.
10. Смешанная фауна.
83. Сравнительная характеристика биоценозов тундры и лесотундры
1. В плакорных условиях и, особенно, на склонах южной экспозиции лесотундры присутствуют острова леса (береза, ель, лиственница) – редины и редколесья. Деревья угнетены, имеют искривленный своеобразный вид ствола и кроны. Тундровые сообщества лесотундры преимущественно пятнистые и кустарниковые. Огромные площади в пределах лесотундры занимают болота.
2. В редколесьях лесотундры живут немногочисленные древесно-кустарниковые виды птиц (варакуша, врановые).
3. Возрастание семенных кормов по сравнению с тундрой объясняет повышенные видовой состав и численность мышевидных грызунов.
4. Ослабление ветров древесным и кустарниковым ярусами способствуют развитию множества летающих насекомых (мухи, комары).
5. Из-за большей мощности деятельного слоя больше почвенных животных (беспозвоночных).
6. В лесотундру зимой откочевывают многие тундровые виды; здесь они находят в снегу укрытия, а под снегом – корм (куропатки, северный олень, полевки).
7. В связи с большим разнообразием фауны в лесотундре устанавливаются более сложные трофические отношения, чем в тундре.
84. Сравнительная характеристика биомов степи и лесостепи
1. Лесостепь характеризуется распространением на плакорах наряду с типичными степными сообществами лесных массивов. Леса занимают пониженные участки в пределах водоразделов.
2. Степные сообщества лесостепи представлены северными разнотравными, или луговыми, степями. Велика роль собственно луговых растений – мезофитов, остальные растения – мезоксерофиты и ксеромезофиты, которые также менее засухоустойчивые, нежели степные. В настоящих степях же преобладают ксерофиты.
3. В лесостепи почти отсутствуют весенные эфемерные растения, распространенные в настоящих степях, особенно в южных.
4. Лесостепи характеризуются большими, чем степи, видовой насыщенностью, густым и высоким травостоем, задернением почвы (здесь оно сплошное); сложной надземной и подземной ярусностью, наличием мохового покрова из мха туидиума.
5. В лесостепи наблюдается максимальное число аспектов.
6. Фауна лесостепи характеризуется обилием лесных видов наряду со степными.